14 January 2021: Lab/In Vitro Research
Mechanical Stress Affects Circadian Rhythm in Skeletal Muscle (C2C12 Myoblasts) by Reducing Per/Cry Gene Expression and Increasing Bmal1 Gene Expression
Mengjia Wang1BCDEF, Da Yu2BCDF, Lichun Zheng1BCDF, Bing Hong1BCD, Houxuan Li1BCD, Xiaobei Hu1BC, Kun Zhang1BF, Yongbin Mou1ADEG*DOI: 10.12659/MSM.928359
Med Sci Monit 2021; 27:e928359






Abstract
BACKGROUND: Circadian rhythm can modulate normal activity of humans in adapting to daily environment changes. Mechanical stress loading affects skeletal muscle development and bio-functions. This study aimed to investigate the effects of mechanical stress loading on circadian rhythm in skeletal muscle (C2C12 cells) and to explore the associated mechanism.
MATERIAL AND METHODS: C2C12 myoblasts were cultured and treated with mechanical stress loading. After mechanical stress loading for 6 h,12 h, and 24 h, we observed the C2C12 myoblasts and determined gene transcription and protein expression of Clock genes, including Clock, Bmal1, Per, and Cry using RT-PCR and western blot assay.
RESULTS: Mechanical stress loading triggered C2C12 cells growing by force direction and enhanced the cell proliferation at 6 h, 12 h, and 24 h. Gene transcription and protein expression of the core Clock-associated molecules, Clock and Bmal1, increased from start of loading to 12 h, and decreased from 12 h to 24 h. Gene transcription and protein expression of core Clock-associated molecules, Cry and Per, decreased in the first 12 h (from 6 h to 12 h) and increased in the last 12 h (from 12 h to 24 h).
CONCLUSIONS: Our study revealed that mechanical stress loading affected circadian rhythm in skeletal muscle (C2C12 myoblasts) through reducing Per/Cry and enhancing Clock/Bmal1 gene expression. This study provides insights for investigating circadian rhythm and associated bio-functions of humans.
Keywords: Circadian clocks, Circadian Rhythm, Myoblasts, Stress, Mechanical, ARNTL Transcription Factors, CLOCK Proteins, Cell Line, Cryptochromes, Period Circadian Proteins
Background
In mammals, circadian rhythms can manipulate fundamental mechanisms, responding to earth cycles [1]. However, many diseases are associated with the abnormality of circadian clocks in human rhythm [2]. Clinically, patients using orthodontic functional appliances at night demonstrate better treatment results than those using such appliances during the day [3,4]. Therefore, greater attention should be focused on the effects of circadian rhythm on muscle function.
Circadian rhythm is regulated by a series of Clock genes, including
Gene transcription for
The
Our clinical experiments showed a difference between the daytime
Previous studies mainly focused on the circadian clock-associated gene expressions in myoblasts; however, the specific mechanism for the effects of mechanical stress loading on myoblasts have been unclear. This study assessed the role of circadian clock-associated genes
Material and Methods
STUDY DESIGN:
We cultured myoblasts and C2C12 cells and subjected them to mechanical stress loading at different timepoints. The proliferation of C2C12 cells undergoing stress loading was examined, and we assessed the circadian clock-associated genes (
This study was approved by the Ethics Committee of the Medical School of Nanjing University, Nanjing, China.
CELL CULTURE AND CHARACTERIZATION OF C2C12 MYOBLASTS:
C2C12 myoblasts were purchased from the China Scientific Institute Cell Center (Shanghai, China). The C2C12 cells were seeded and then cultured in DMEM (Gibco BRL. Co., Ltd., Grand Island, NY, USA), supplemented with 10% FBS (Gibco BRL. Co. Ltd.) containing 1% penicillin-streptomycin (Sigma-Aldrich, St. Louis, MO, USA), at 37°C and 5% CO2. The culture medium was replaced every 2–3 days, keeping cells with 80% cell fusion.
GENERATION FOR CYCLIC MECHANICAL STRESS MODEL:
C2C12 myoblasts were seeded into a culture bottle and cultured, and then observed under a microscope. The old culturing medium in the culture bottle was removed, followed by washing 2–3 times with PBS (Beyotime Biotech, Shanghai, China). About 50 ml culture medium containing 1–3 ml 0.25% trypsin solution (Beyotime Biotech) was added into the culture bottle. The culture bottle was shaken to evenly distribute the digestive juices, and the cells were then incubated at 37°C for 2–5 min. Under the microscope, when the C2C12 cells shrank or started falling off, we added 2–3 ml complete culturing medium followed by blowing gently, and the C2C12 myoblasts were collected into a centrifuge tube. Then, the C2C12 myoblasts were centrifuged to obtain the cell pellets and we discarded the supernatant. The cell pellets were dissolved into the DMEM medium and adjusted to a density of 6×104 cells/ml.
When the cells were fused to about 80%, the serum-free culture group was replaced for further culture for 24 h to synchronize the cells. Then, the post-conditioning treatment was started. In this study, mechanical stress was loaded as described in our former study [14]. Briefly, C2C12 myoblasts received application of 15% mechanical stress loading and 0.5 Hz for 0 h, 6 h, 12 h, and 24 h using a Flexercell FX-4000 strain unit (Flexcell Int. Co., McKeesport, PA, USA). The loading stress was conducted at room temperature with 10 cycles/min, and each cycle included 3 s of stretch and 3 s of relaxation.
TRIAL GROUPING:
C2C12 myoblasts were divided into 4 groups: a Normal cell group, a Stress 6 h group, a Stress 12 h group, and a Stress 24 h group, with 6 samples for each group (n=6). The Normal cell group was assigned as the normal C2C12 myoblasts without loading stress. The Stress 6 h group was assigned as C2C12 myoblasts loading with 15% mechanical stress for 6 h. The Stress 12 h group was assigned as C2C12 myoblasts loading with 15% mechanical stress for 12 h. The Stress 24 h group was assigned as C2C12 myoblasts loading with 15% mechanical stress for 24 h. For all groups, the cell amounts refer to the total number of cells in the culture bottle. All of the experiments were repeated at least 6 times.
WESTERN BLOT ASSSAY:
The suspended C2C12 myoblasts were collected, centrifuged, and washed once with PBS. The cells were treated with RIPA buffer (Beyotime Biotech) on ice for 20 min to treat cells. Then, the lysates were added to a 1.5-ml centrifuge tube and strongly agitated for 30 s. After centrifuging at 12 000 rpm for 15 min at 4°C, the supernatant was collected for subsequent western blot assay. Then, cell lysates were loaded onto 12% SDS-PAGE (Beyotime Biotech) and electro-transferred onto the PVDF membranes (Beyotime Biotech). PVDF membranes were incubated with anti-mouse Per, anti-mouse Cry, anti-mouse Clock, anti-mouse Bmal1, and anti-mouse β-actin overnight at 4°C. PVDF membranes were subsequently incubated with HRP-conjugated IgG at room temperature for 2 h. All of the above primary and secondary antibodies were purchased from Abcam Biotech (Cambridge, MA, USA). The western blot bands were visualized using the BeyoECL Plus Kit (Cat. No. P0018S, Beyotime Biotech) according to the manufacturer’s instructions. The staining images were captured using a professional imaging system (ProteinSimple, Santa Clara, CA, USA).
RT-PCR ASSAY:
The RT-PCR assay was carried out to determine gene transcriptions for the Per gene, Cry gene, Clock gene, and Bmal1 gene. The total RNAs were extracted with Trizol reagents and then reversed to synthesize the cDNAs for conducting the RT-PCR assay. The primers for Per, Cry, Clock, Bmal1, and the internal reference (GAPDH) were designed and synthesized as illustrated in Table 1. The RT-PCR assay conditions were listed as the followings: initial denaturation at 94°C for 4 min, followed by 35 cycles at 94°C 30 s, 60°C for 30 s and 72°C for 45 min, with a final cycle at 72°C for 10 min The transcriptions for the above targeted genes were normalized to GAPDH. The relative gene transcription for the gene was calculated as described by a previous report using the 2−ΔΔCt method [19].
STATISTICAL ANALYSES:
Data are represented as mean±standard deviation (SD) and analyzed using the professional statistical software package SPSS (version: 18.0, SPSS Inc, Chicago, IL, USA). Tukey’s post hoc test-validated ANOVA was utilized for comparing differences of measurement data among groups. Statistical significance was defined when the P value was less than 0.05.
Results
MECHANICAL STRESS PROMOTED C2C12 PROLIFERATION:
In this study, the growth status of C2C12 myoblasts undergoing mechanical stress loading were observed first. C2C12 myoblasts in the Normal group were cultured and distributed as represented in Figure 1A. After loading with mechanical stress on C2C12 cells, cells were arranged and separated at 6 h (Figure 1B), 12 h (Figure 1C), and 24 h (Figure 1D) and compared to those in the Normal group. Moreover, the cells loaded with stress for 12 h and 24 h conformed to the loading direction (Figure 1C, 1D).
Because extra-cellular stress can influence cell growth, we observed the status of C2C12 myoblasts with a microscope. Cell counts of each group were compared with cell counts in the Normal group. We observed a distinct difference in stress-loaded cells, including cells distributing with the force direction and accumulating in clusters (Figure 1A). When treating cells with mechanical stress for 6 h, the proliferation of myoblasts was promoted (Figure 1B). When cells were treated with cyclic tension for 12 h, the number of cells was increased dramatically and cells were densely clustered (Figure 1C). However, there was a little cell death in cells undergoing stress for 12 h, because C2C12 cells showed nuclear chromatin condensation and fragmentation (Figure 1C). Following exposure to stress loading for 24 h, C2C12 myoblasts were assembled and a few dead cells appeared (Figure 1D). The cell numbers peaked at 12 h after stress loading (Figure 1E).
:
The gene transcriptions for Per and Cry were determined using RT-PCR assay and western blot assay. The results demonstrated that mechanical stress significantly decreased the Per gene (Figure 2A) and Cry gene (Figure 2B) transcription compared to those at 12 h after stress loading (P<0.05). However, the Per gene (Figure 2A) and Cry gene (Figure 2B) transcriptions were remarkably increased at 24 h after stress loading compared to those at 12 h after stress loading (P<0.001). Moreover, the western blot findings (Figure 2C) also identified that Per (Figure 2D) and Cry (Figure 2E) expressions in cells were significantly decreased at 12 h after stress loading compared to those in the Normal group (P<0.01). However, Per (Figure 2D) and Cry (Figure 2E) expressions were significantly increased at 24 h after stress loading compared to those at 12 h after loading stress (P<0.001). These results suggest that Per and Cry transcriptions and expressions may be inhibited following mechanical stress loading, and the expressions were regenerated with the accumulation of stress.
:
We found that transcriptions for the Clock gene (Figure 3A) and Bmal1 gene (Figure 3B) were markedly increased at 12 h after stress loading compared to those in the Normal cells (P<0.05). Clock gene (Figure 3A) and Bmal1 gene (Figure 3B) transcriptions at 24 h after stress loading were significantly decreased compared to those at 12 h after loading stress (P<0.001).
Furthermore, western blot results (Figure 3C) indicated that mechanical stress (12 h loading) significantly increased the Clock (Figure 3D) and Bmal1 (Figure 3E) expression compared to those in Normal cells (P<0.05). However, the mechanical stress (24 h loading) remarkably reduced the Clock (Figure 3D) and Bmal1 (Figure 3E) expression compared to those at 12 h after stress loading (P<0.001). In summary, these results suggest that mechanical stress promotes Clock and Bmal1 (6 h and 12 h loading stress) and then induced apoptosis over time (24 h loading stress).
:
According to the gene transcription potential of Per, Cry, Clock, and Bmal1 molecules in C2C12 cells undergoing mechanical stress loading from 6 h to 24 h, we found that the appropriate timepoint for the Per (Figure 4A), Cry (Figure 4B), Clock (Figure 4C), and Bmal1 (Figure 4D) expression is 12 h after mechanical stress loading. The gene transcription of Per (Figure 4A) and Cry (Figure 4B) at 12 h after mechanical stress loading was remarkably lower compared to those at 6 h and 24 h after mechanical stress loading (P<0.05). Meanwhile, the gene transcription of Clock (Figure 4C) and Bmal1 (Figure 4D) at 12 h after mechanical stress loading was markedly higher compared to those at 6 h and 24 h after mechanical stress loading (P<0.05). These results suggest that short (12 h) mechanical stress loading can reduce Per/Cry and enhance Clock/Bmal1 expression, but long-term (24 h) loading can suppress these effects.
Discussion
The present study demonstrated that mechanical stress loading triggered C2C12 myoblasts growth by force direction and enhanced the cell proliferation. The gene transcription and expression of core Clock-associated molecules,
Life on earth has adapted to the environment throughout its long evolutionary history. The periodic changes in its physiological activities form a variety of different time cycles, which include biological rhythm (circadian clock). The circadian clock is a self-regulated system which is generated from self-sustaining oscillator composition [20]. The synchronization is not only the body clock cycle and the rotation of the earth, but, more importantly, also the stable relation between time and phase [21,22].
In mammals, the circadian clock, as an oscillation, is found in almost all cells and tissues of the body. The circadian clock also participates in optimization and regulation of physiological activities of tissues or organs. Rosbash et al. [23] discovered the first circadian gene in fruit flies, called Period. The transcription products of circadian gene showed a rhythm change of approximately 24 h [24,25]. Previous studies [26,27] continued the research and used biochemical methods, and then found the Clock gene.
Vitaterna et al. [28] found that the function of the
However, skeletal muscle molecular clocks depend not only on environmental factors, but also on synchronization with other tissues. When the circadian rhythm of the human body is interrupted, skeletal muscle usually undergoes a series of changes, including myofibril structure, fiber optic transformation, damaged muscle functions, and reduced mitochondrial respiration. Additionally, there are many adverse effects or side-effects on the metabolism when the circadian rhythm is interrupted, such as insulin sensitivity and glucose tolerance. Therefore, skeletal muscle might be an important metabolism-related tissue. The above background demonstrates that circadian rhythm in skeletal muscle plays critical roles in muscle tissues and even the whole body. Brager et al. [31] found that the Bmal1 gene was overexpressed in bone and muscle, and could affect the cell metabolism process Ping et al. [32] also reported differences in Clock genes in fast and slow muscle fibers. The difference in Clock gene expression of muscle fibers also identifies the critical functions of the circadian system in maintaining biological functions of muscle tissues. Therefore, further research is necessary to explore the specific mechanism of the biological clock in skeletal muscle and to evaluate the expression of circadian rhythm-associated genes in skeletal muscles.
Schiaffion et al. [33] established the Bmal1 knock-out model, which explained muscle metabolism. The change comes from interruption of the muscle clock rather than the special function of BMAL1. Clock genes mainly exist in muscle, bone, cartilage, and tendon tissues. A previous study [34] focused on circadian rhythms in tissue physiology and disease-associated tissues [34]. More advanced research will help understand how circadian rhythms control muscle and bone health and prevent musculoskeletal tissue disease, as well as showing how to improve the efficacy of orthodontic functional appliances in the orthodontic treatment process. To explore biological characteristics of circadian genes in myoblasts, intermittent cyclic stress was selected as an experimental strategy. In the present study, cyclic mechanical stress at 15% elongation with 0.5 Hz was used to simulate oral biomechanical stimulation on C2C12 myoblasts.
There are a series of factors involved in myoblast proliferation, such as the mechanical stress. Our recent studies indicated that constant stress promotes protein expression and mRNA synthesis at the beginning of stretching. Therefore, we detected cell accumulation and growth for the first 12 h of a day and rarely observed nucleus fragmentation. When the stress lasted for a whole day, the proliferation began to decline and more nucleus fragmentation was found. Our results conclude that the mechanical stress reaches to its maximum in the first half of the day, which means the orthodontic functional appliance should be worn for 12 h during the first cycle of the day without being taken off.
The gene transcriptions of circadian genes, including
Andrew et al. [18] discovered that MyoD1, which is a well-characterized molecule, regulate the transcriptions of muscle genes under the direct control of the Bmal1/Clock signaling pathway. Chatterjee et al. [35] also reported that Bmal1 could temporally modulate myogenic differentiation. Myogenin, a transcriptional factor inducing myogenesis, has been proven to be a circadian molecule [36]. The above studies demonstrated critical interactions between molecular clock mechanism and associated transcriptional factors such as MyoD1 and Myogenin. The phase of the molecular clock can be identified by the time of peak
Mcdearmon et al. [38] reported an important function of the skeletal muscle clock in maintaining skeletal muscle health and systemic health. We also found in normal cells the expression of Bmal1 was lower than the mechanical stress loading. The results indicated that mechanical stress promotes skeletal muscle health and increased the circadian clock rhythm, suggesting orthodontic functional appliances should be worn as much as possible.
Circadian rhythms have been proven to be oscillations of about 24 h and can modulate the transcription and metabolism of genes in muscles. Per, as a crucial upstream factor for the p53 signaling pathway, plays critical roles in modulating the proliferation of cells, cell apoptosis, and cell cycles. Furthermore, Per can inhibit tumor growth and repair DNA damage. The present results show that Per transcription was decreased in the first cycle for 12 h under stress and was increased in the later cycle. According to our data, mechanical stress inhibits Per expression compared with the normal cells, and after loading more than half of a day, it will promote Per, which increases proliferation.
A recent study [39] has reported that Cry gene variation leads to late sleeping. Furthermore, the low expression of Cry will lead the interruption of normal circadian rhythm. In our research, we have found that Cry expression decreased during the first cycle, which means mechanical stress could not promote Cry during the first half of the day. The results show that the low expression of Cry will interrupt the normal cycle. A previous study [39] demonstrated that disruptions of circadian rhythms can damage functions of skeletal muscles. Therefore, we can conclude that the mechanical stress should be continued for 12 h.
From the above discussion, our data first suggest that the orthodontic functional appliance should be wore for at least 12 h. Moreover, the light could enhance the Clock/Bmal1 expression and promote the myoblasts growth. While, after lasting for more than 12 h of light, the Per and Cry could also be activated and increase the myoblasts proliferation. As a result, functional applicant should be used for more than 12 h light in one day. Second, our data also showed that, compared with nighttime only or daytime only wearing, wearing the orthodontic functional appliance all day exerts the strongest effect on myoblasts. Based on a circadian cycle of 24 h, we have found that Clock/Bmal1 and Per/Cry play opposite roles in the whole cycle. Therefore, both of Clock/Bmal1 and Per/Cry are consist of the oscillation period. We suggest that patients wear the orthodontic functional appliance for the whole day. Third, Bmal1 plays an important role in skeletal muscle rhythm and is highly expressed during the daytime due to the light cue. If appliance wearing time is limited, we recommend wearing the orthodontic functional appliance during the daytime.
Many studies [13,14,40–42] have reported associations between mechanical stress loading and proliferation of C2C12 myoblasts. Vitale et al. [43] reviewed previous studies and concluded that circadian rhythm in skeletal muscle is crucial for maintaining cellular metabolism, physiology, and bio-functions. However, the present study investigated effects of mechanical stress loading on proliferation of circadian rhythm in skeletal muscle for the first time. This study assessed the role of circadian rhythm-associated genes
Although some interesting results were discovered, there are also a few limitations. First, the effects of mechanical stress on the improvement of muscle rhythmicity were not determined in this study. Second, the myogenic regulatory factors (MRFs) modulating skeletal muscle, such as myoD, were not examined in mechanical stress-loaded C2C12 myoblasts. The identification of effects of mechanical stress on MRFs prove that mechanical force can indeed promote muscle development. Third, as our results indicated, circadian rhythm-associated genes expression in C2C12 myoblasts under mechanical stress loading might reflect rhythmic expression, but different gene expression in C2C12 myoblasts undergoing stress loading and without stress treatment were not compared. Fourth, this study clarified effects of mechanical stress loading, but the relationship between rhythm gene expression and C2C12 myoblasts proliferation was not studied.
Conclusions
The present study demonstrates that mechanical stress loading affects circadian rhythm in skeletal muscle (C2C12 myoblasts) through modulating circadian rhythm-associated molecules (reducing Per/Cry and enhancing Clock/
Figures
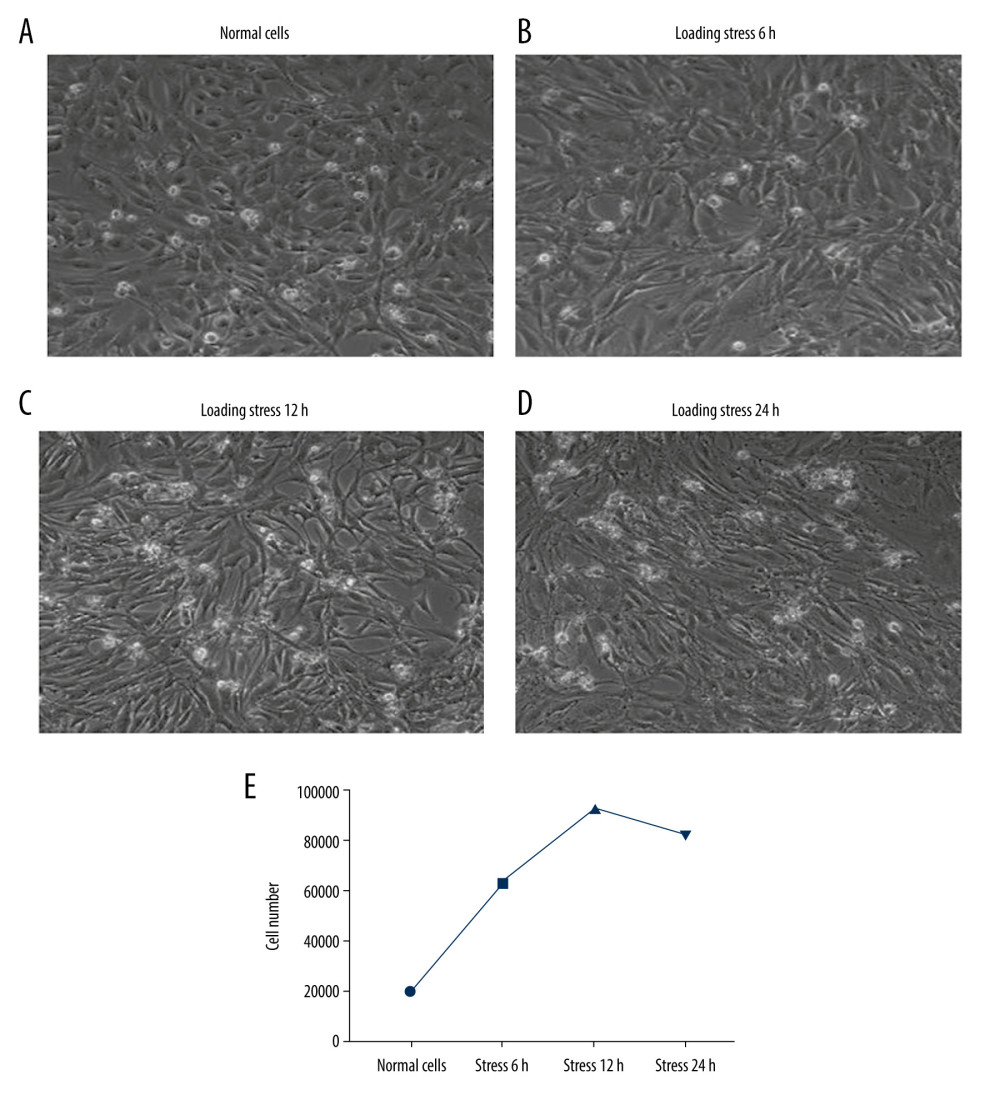 Figure 1. Characteristics for the C2C12 myoblasts. (A) Normal cells (n=6). (B) Cells loaded with stress for 6 h (n=6). (C) Cells loaded with stress for 12h (n=6). (D) Cells loaded with stress for 24 h (n=6). (E) Cell amounts in Normal cells and cells undergoing different stress loading.
Figure 1. Characteristics for the C2C12 myoblasts. (A) Normal cells (n=6). (B) Cells loaded with stress for 6 h (n=6). (C) Cells loaded with stress for 12h (n=6). (D) Cells loaded with stress for 24 h (n=6). (E) Cell amounts in Normal cells and cells undergoing different stress loading. 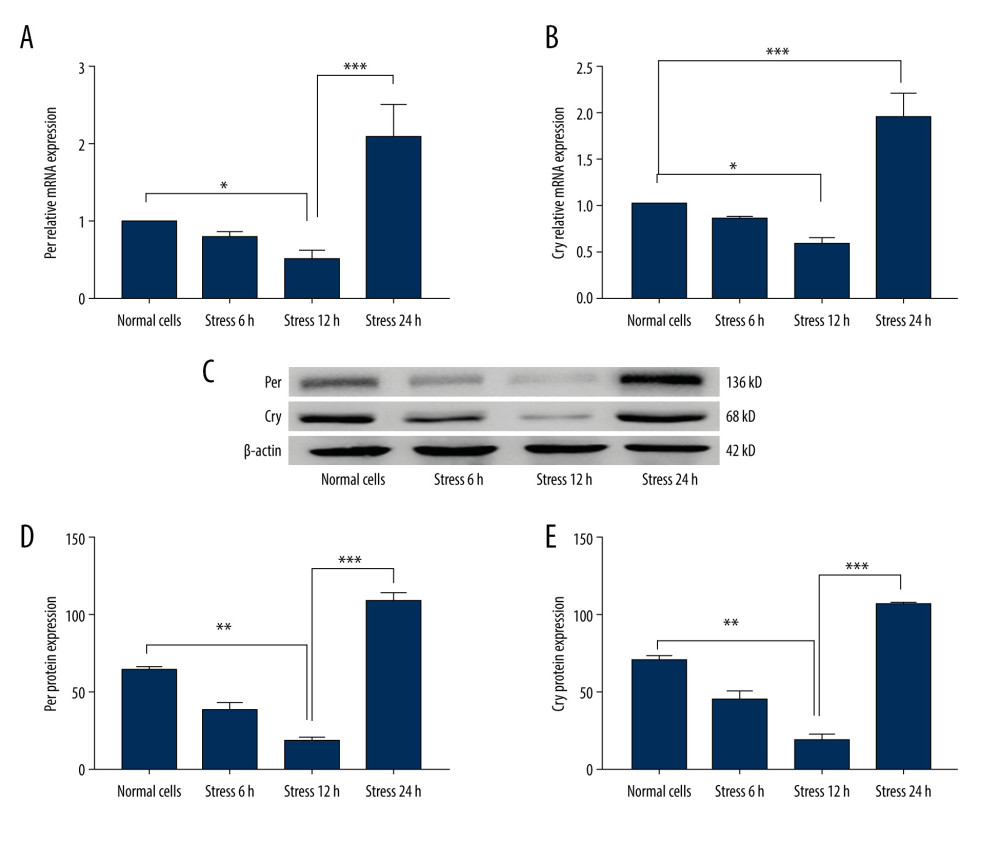 Figure 2. Evaluation for the Per and Cry gene transcriptions and protein expressions in C2C12 myoblasts (n=6). (A) Statistical analysis for the gene transcription of Per determining by PCR assay. (B) Statistical analysis for the gene transcription of Cry determining by PCR assay. (C) Western blot images for the Per and Cry protein expression. (D) Statistical analysis for the protein expression of Per in C2C12 cells. (E) Statistical analysis for the protein expression of Cry in C2C12 cells. * P<0.05, ** P<0.01, and *** P<0.001 represents the differences illustrated in images.
Figure 2. Evaluation for the Per and Cry gene transcriptions and protein expressions in C2C12 myoblasts (n=6). (A) Statistical analysis for the gene transcription of Per determining by PCR assay. (B) Statistical analysis for the gene transcription of Cry determining by PCR assay. (C) Western blot images for the Per and Cry protein expression. (D) Statistical analysis for the protein expression of Per in C2C12 cells. (E) Statistical analysis for the protein expression of Cry in C2C12 cells. * P<0.05, ** P<0.01, and *** P<0.001 represents the differences illustrated in images. 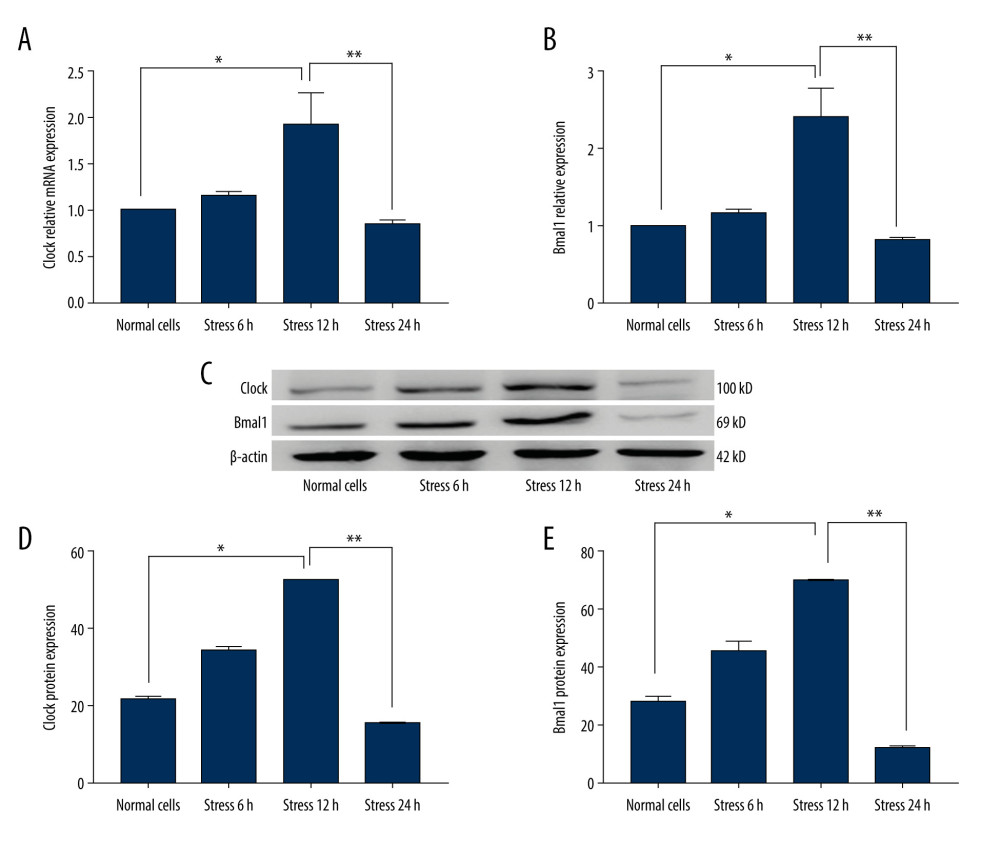 Figure 3. Determination of the Per and Cry gene transcriptions and protein expressions in C2C12 myoblasts (n=6). (A) Statistical analysis for Clock gene transcription identified by PCR assay. (B) Statistical analysis for Bmal1 gene transcription of Cry identified by PCR assay. (C) Western blot images for Clock and Bmal1 protein expression. (D) Statistical analysis for Clock expression in C2C12 cells. (E) Statistical analysis for Bmal1 expression in C2C12 cells. * P<0.05 and ** P<0.001 represent differences illustrated in images.
Figure 3. Determination of the Per and Cry gene transcriptions and protein expressions in C2C12 myoblasts (n=6). (A) Statistical analysis for Clock gene transcription identified by PCR assay. (B) Statistical analysis for Bmal1 gene transcription of Cry identified by PCR assay. (C) Western blot images for Clock and Bmal1 protein expression. (D) Statistical analysis for Clock expression in C2C12 cells. (E) Statistical analysis for Bmal1 expression in C2C12 cells. * P<0.05 and ** P<0.001 represent differences illustrated in images. 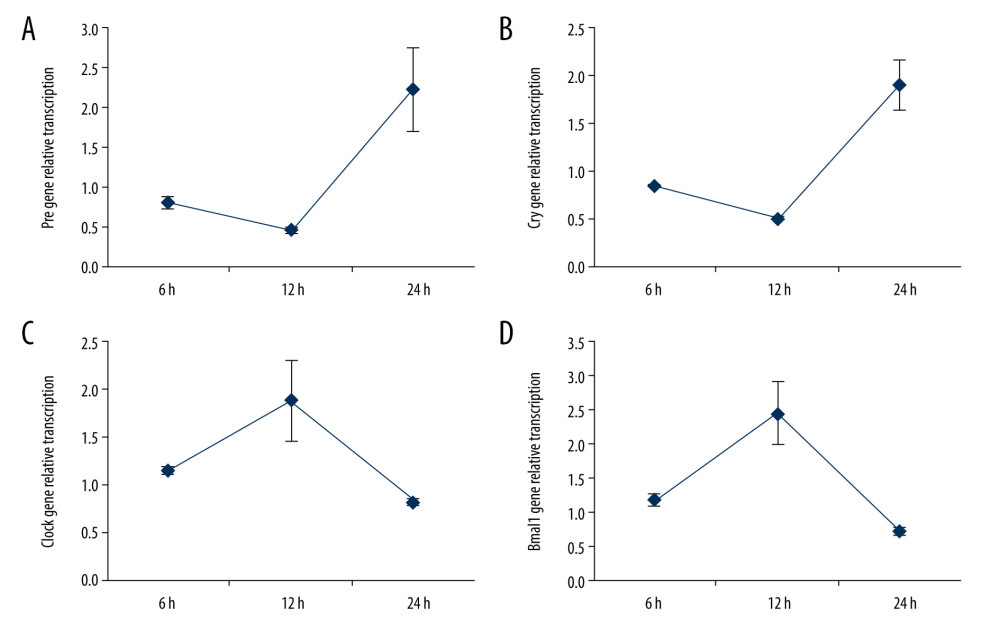 Figure 4. Determinations for the gene transcription tendency of Per (A), Cry (B), Clock (C), and Baml1 (D) in C2C12 myoblasts from 6 h to 24 h. The gene transcriptions were determined with PCR assay (n=6).
Figure 4. Determinations for the gene transcription tendency of Per (A), Cry (B), Clock (C), and Baml1 (D) in C2C12 myoblasts from 6 h to 24 h. The gene transcriptions were determined with PCR assay (n=6). 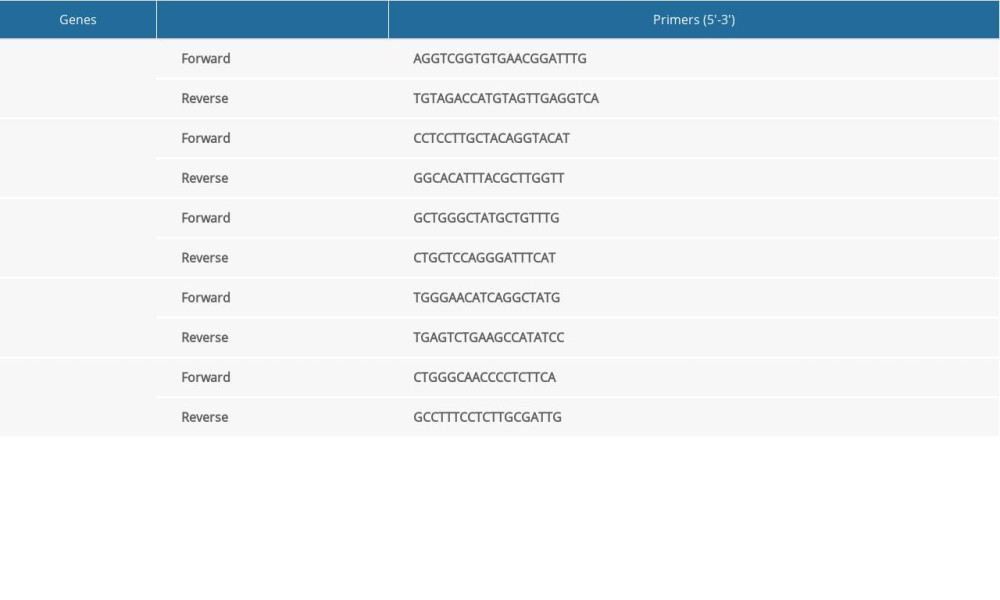
References
1. Huang W, Ramsey KM, Marcheva B, Circadian rhythms, sleep, and metabolism: J Clin Invest, 2011; 121; 2133-41
2. Gabriel BM, Zierath JR, Circadian rhythms and excise, re-setting the clock in metabolic disease: Nat Rev Endocrinol, 2019; 15; 197-206
3. Sander FG, Functional processes when wearing the SII appliance during the day: J Orofac Orthop, 2001; 62; 264-74
4. Celli D, Gasperoni E, Oliva B, Assessment of mandibular growth and response to functional appliance treatment in prepubertal patients with different auxologic categories: Prog Orthod, 2012; 11; 20-26
5. Jagannath A, Taylor I, Wakaf Z, The genetics of circadian rhythms, sleep and health: Hum Mol Genet, 2017; 26; R128-38
6. Lamont EW, James FO, Boivin DB, From circadian clock gene expression to pathologies: Sleep Med, 2007; 8; 547-56
7. Lee J, Lee S, Chung S, Identification of a novel circadian clock modulator controlling BMAL1 expression through a ROR/REV-ERB-response element-dependent mechanism: Biochem Biphys Res Commun, 2016; 469; 580-86
8. Wang Z, Wu Y, Li L, Intermolecular recognition revealed by the complex structure of human CLOCK-BMAL1 basic helix-loop-helix domains with E-box DNA: Cell Res, 2013; 23; 213-24
9. Guillaumond F, Dardente H, Giguere V: J Biol Rhythms, 2005; 20; 391-403
10. Hirayama J, Sahar S, Grimaldi B, CLOCK-mediated acetylation of BMAL1 controls circadian function: Nature, 2007; 450; 1086-90
11. Kim TJ, Sung JH, Shin JC, CRISPR/Cas-mediated Fubp1 silencing disrupts circadian oscillation of Per1 protein via downregulating Syncrip expression: Cell Biol Int, 2020; 44; 424-32
12. Ebling FJ, The role of glutamate in the photic regulation of the suprachiasmatic nucleus: Prog Neurobiol, 1996; 50; 109-32
13. Wang M, Mou Y, Da Y, Effects of mammalian target of rapamycin on proliferation, apoptosis and differentiation of myoblasts undergoing mechanical stress: Am J Transl Res, 2018; 10; 4173-82
14. Da Y, Mou Y, Wang M, Mechanical stress promotes biological functions of C2C12 myoblasts by activating PI3K/AKT/mTOR signaling pathway: Mol Med Rep, 2020; 21; 470-77
15. Birbrair A, Zhang T, Wang ZM, Role of pericytes in skeletal muscle regeneration and fat accumulation: Stem Cells Dev, 2013; 22; 2298-314
16. Knoll WD, Gaida A, Maurer P, Analysis of mechanical stress in reconstruction for bridging mandibular angle defects: J Craniomaxillofac Surg, 2006; 34; 201-9
17. Harfmann BD, Schroder EA, Esser KA, Circadian rhythms, the molecular clock, and skeletal muscle: J Biol Rhythms, 2015; 30; 84-94
18. Andrews JL, Zhang X, McCarthy JJ, CLOCK and BMAL1 regulate MyoD and are necessary for maintenance of skeletal muscle phenotype and function: Proc Natl Acad Sci USA, 2010; 107; 19090-95
19. Livak KJ, Schmittgen TD: Methods, 2001; 25; 402-8
20. Kondratova AA, Kondratov RV, The circadian clock and pathology of the ageing brain: Nat Rev Neurosci, 2012; 13; 325-35
21. Duffy JF, Wright KP, Entrainment of the human circadian system by light: J Biol Rhythms, 2005; 20; 326-38
22. Qin XM, Guo JH, Synchronization of the mammalian central and peripheral circadian clocks: Chin Sci Bull, 2017; 62; 2849-56
23. Rosbash M, Allada R, McDonald M, Circadian rhythms in Drosophila: Novartis Found Symp, 2003; 253; 223-32
24. Reddy P, Zehring WA, Wheeler DA, Molecular analysis of the period locus in Drosophila melanogaster and identification of a transcript involved in biological rhythms: Cell, 1984; 38; 701-10
25. Bargiello TA, Jackson FR, Young MW, Restoration of circadian behavioural rhythms by gene transfer in Drosophila: Nature, 1984; 312; 752-54
26. Allada R, White NE, So WV, A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless: Cell, 1998; 93; 791-804
27. Rutila JE, Suri V, Le M, CYCLE is a second bHLH-PAS clock protein essential for circadian rhythmicity and transcription of Drosophila period and timeless: Cell, 1998; 93; 805-14
28. Vitaterna MH, King DP, Chang AM, Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior: Science, 1994; 264; 719-25
29. Gekakis N, Staknis D, Nguyen HB, Role of the CLOCK protein in the mammalian circadian mechanism: Science, 1998; 280; 1564-69
30. Bunger MK, Wilsbacher LD, Moran SM, Mop3 is an essential component of the master circadian pacemaker in mammals: Cell, 2000; 103; 1009-17
31. Brager AJ, Heemstra L, Bhambra R, Homeostatic effects of exercise and sleep on metabolic processes in mice with an overexpressed skeletal muscle clock: Biochimie, 2017; 132; 161-65
32. Wu Ping, Li YL, Cheng J, Daily rhythmicity of clock gene transcript levels in fast and slow muscle fibers from Chinese perch (Siniperca chuatsi): BMC Genomic, 2016; 17; 1008
33. Schiaffino S, Blaauw B, Dyar KA: Skeletal Muscle, 2016; 6; 33
34. Dudek M, Meng QJ, Running on time: The role of circadian clocks in the musculoskeletal system: Biochem J, 2014; 463; 1-8
35. Andrews JL, Zhang X, McCarthy JJ, CLOCK and BMAL1 regulate MyoD and are necessary for maintenance of skeletal muscle phenotype and function: Proc Natl Acad Sci USA, 2010; 107; 19090-95
36. Chatterjee S, Nam D, Guo B, Brain and muscle Arnt-like 1 is a key regulator of myogenesis: J Cell Sci, 2013; 126; 2213-24
37. Shavlakadze T, Anwari T, Soffe Z, Impact of fasting on the rhythmic expression of myogenic and metabolic factors in skeletal muscle of adult mice: Am J Physiol Cell Physiol, 2013; 305; C26-35
38. Roenneberg T, Daan S, Merrow M, The art of entrainment: J Biol Rhythms, 2003; 18; 183-94
39. McDearmon EL, Patel KN, Ko CH, Dissecting the functions of the mammalian clock protein BMAL1 by tissue-specific rescue in mice: Science, 2006; 314; 1304-8
40. Yoo SH, Yamazaki S, Lowrey PL, PERIOD2: LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues: Proc Natl Acad Sci USA, 2004; 101; 5339-46
41. Matheny RW, Adamo ML, Effects of PI3K catalytic subunit and Akt isoform deficiency on mTOR and p70S6K activation in myoblasts: Biochem Biophys Res Commum, 2009; 390; 252-57
42. Hu SY, Tai CC, Li YH, Progranulin compensates for blocked IGF-1 signaling to promote myotube hypertrophy in C2C12 myoblasts via the PI3K/Akt/mTOR pathway: FEBS Lett, 2012; 586; 3485-92
43. Kitakaze T, Sakamoto T, Kitano T, The collagen derived dipeptide hydroxyprolyl-glycine promotes C2C12 myoblast differentiation and myotube hypertrophy: Biochem Biophys Res Commun, 2016; 478; 1292-97
44. Vitale JA, Bonato M, La Torre A, The role of the molecular clock in promoting skeletal muscle growth and protecting against sarcopenia: Int J Mol Sci, 2019; 20; 4318
Figures
Figure 1. Characteristics for the C2C12 myoblasts. (A) Normal cells (n=6). (B) Cells loaded with stress for 6 h (n=6). (C) Cells loaded with stress for 12h (n=6). (D) Cells loaded with stress for 24 h (n=6). (E) Cell amounts in Normal cells and cells undergoing different stress loading.Figure 2. Evaluation for the Per and Cry gene transcriptions and protein expressions in C2C12 myoblasts (n=6). (A) Statistical analysis for the gene transcription of Per determining by PCR assay. (B) Statistical analysis for the gene transcription of Cry determining by PCR assay. (C) Western blot images for the Per and Cry protein expression. (D) Statistical analysis for the protein expression of Per in C2C12 cells. (E) Statistical analysis for the protein expression of Cry in C2C12 cells. * P<0.05, ** P<0.01, and *** P<0.001 represents the differences illustrated in images.Figure 3. Determination of the Per and Cry gene transcriptions and protein expressions in C2C12 myoblasts (n=6). (A) Statistical analysis for Clock gene transcription identified by PCR assay. (B) Statistical analysis for Bmal1 gene transcription of Cry identified by PCR assay. (C) Western blot images for Clock and Bmal1 protein expression. (D) Statistical analysis for Clock expression in C2C12 cells. (E) Statistical analysis for Bmal1 expression in C2C12 cells. * P<0.05 and ** P<0.001 represent differences illustrated in images.Figure 4. Determinations for the gene transcription tendency of Per (A), Cry (B), Clock (C), and Baml1 (D) in C2C12 myoblasts from 6 h to 24 h. The gene transcriptions were determined with PCR assay (n=6). In Press
07 Mar 2024 : Clinical Research
Knowledge of and Attitudes Toward Clinical Trials: A Questionnaire-Based Study of 179 Male Third- and Fourt...Med Sci Monit In Press; DOI: 10.12659/MSM.943468
08 Mar 2024 : Animal Research
Modification of Experimental Model of Necrotizing Enterocolitis (NEC) in Rat Pups by Single Exposure to Hyp...Med Sci Monit In Press; DOI: 10.12659/MSM.943443
18 Apr 2024 : Clinical Research
Comparative Analysis of Open and Closed Sphincterotomy for the Treatment of Chronic Anal Fissure: Safety an...Med Sci Monit In Press; DOI: 10.12659/MSM.944127
08 Mar 2024 : Laboratory Research
Evaluation of Retentive Strength of 50 Endodontically-Treated Single-Rooted Mandibular Second Premolars Res...Med Sci Monit In Press; DOI: 10.12659/MSM.944110
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952