19 January 2021: Database Analysis
Bioinformatics Analysis of Differentially Expressed Genes and Protein–Protein Interaction Networks Associated with Functional Pathways in Ulcerative Colitis
Feng Cao1AE, Yun-Sheng Cheng1E, Liang Yu1D, Yan-Yan Xu2AG*, Yong Wang1ADOI: 10.12659/MSM.927917
Med Sci Monit 2021; 27:e927917






Abstract
BACKGROUND: This bioinformatics study aimed to identify differentially expressed genes (DEGs) and protein–protein interaction (PPI) networks associated with functional pathways in ulcerative colitis based on 3 Gene Expression Omnibus (GEO) datasets.
MATERIAL AND METHODS: The GSE87466, GSE75214, and GSE48958 MINiML formatted family files were downloaded from the GEO database. DEGs were identified from the 3 datasets, and volcano maps and heat maps were drawn after R language standardization and analysis, respectively. Venn diagram software was used to identify common DEGs. PPI analysis of common DEGs was performed using the Search Tool for the Retrieval of Interacting Genes. Gene modules and hub genes were visualized in the PPI network using Cytoscape. Enrichment analysis was performed for all common DEGs, module genes, and hub genes.
RESULTS: A total of 90 DEGs were selected, which included 3 functional modules and 1 hub gene module. CXCL8 module genes were mainly enriched in cytokine-mediated signaling pathways and interleukin (IL)-10 signaling. CCL20 module genes were mainly enriched in the IL-17 signaling pathway and cellular response to IL-1. Hub gene modules mainly involved IL-10, IL-4, and IL-13 signaling pathways. CXCL8, CXCL1, and IL-1β were the top 3 hub genes and were mainly involved in IL-10 signaling.
CONCLUSIONS: Bioinformatics analysis using 3 GEO datasets identified CXCL8, CXCL1, and IL-1β, which are involved in IL-10 signaling, as the top 3 hub genes in ulcerative colitis. The findings from this study remain to be validated, but they may contribute to the further understanding of the pathogenesis of ulcerative colitis.
Keywords: Colitis, Ulcerative, Colon, Chemokine CXCL1, Computational Biology, Gene Regulatory Networks, Interleukin-8, Protein Interaction Maps
Background
Inflammatory bowel disease (IBD) includes Crohn’s disease and ulcerative colitis (UC), which have overlapping clinical and molecular features. The incidence rate of IBD is increasing in worldwide. China has the highest incidence of IBD in Asia, and the ratio of UC to Crohn’s disease was reported to be 2.0 [1]. UC is a diffuse, chronic, nonspecific inflammatory disease that occurs in the colorectal mucosa and/or submucosa. The clinical symptoms of UC are characterized by relapsing mucosal inflammation that manifests as persistent or repeated mucopurulent bloody stool, abdominal pain, and other systemic symptoms [2]. UC prevalence is highest in Europe, followed by Canada and then the United States [3]. The age of onset is getting younger [4]. UC is recurrent and difficult to heal, and it is referred to as “deathless cancer” due to its serious impact on patients’ quality of life. UC is closely related to the development of colon cancer [5], which is difficult to cure. The pathogenesis of UC has not been clarified, and the possible underlying mechanisms include bacterial infection, intestinal mucosal barrier dysfunction, and genetic, dietary, environmental, immunological, and psychological factors [6]. The interaction of a variety of factors results in high intestinal mucosa sensitivity, ultimately leading to intestinal inflammation and tissue damage. Conservative medical treatment of UC in clinical practice is somewhat limited and includes dietary regulation and drug treatment with aminosalicylic acid, antibiotics, probiotics, and glucocorticoids. The criterion standard surgical treatment of UC is ileal pouch anal anastomosis [7]. This surgery can remove UC target organs, but the incidence of postoperative complication of pouchitis is high [8,9]. Therefore, further understanding of the pathogenesis and regulation of UC at the molecular level may provide new directions for UC prevention and treatment.
The rapid development and extensive application of gene expression profile data has led to bioinformatics analysis becoming a popular method to explore disease pathogenesis. Bioinformatics analysis provides significant insight into the pathophysiological mechanisms of diseases at the genetic level. For example, Cheng et al. [10] screened GNG11, GNB4, and AGT as potential molecular targets and diagnostic biomarkers for UC and Crohn’s disease by bioinformatics. Feng et al. [11] found that TATAbinding protein 1, nuclear factor-κB, and microRNAs were closely related to 233 differentially expressed genes (DEGs) of UC and may be potential molecular targets for its treatment [11]. Here, using 3 datasets from the Gene Expression Omnibus (GEO) database, we applied bioinformatics tools to perform a series of UC expression profile analyses to identify potential core genes that can serve as molecular targets for the prevention and treatment of UC. Through these analyses, we identified DEGs and protein–protein interaction (PPI) networks associated with functional pathways in UC.
Material and Methods
DATASET INFORMATION:
The search terms used in the GEO database, which is affiliated with the National Center for Biotechnology Information (https://www.ncbi.nlm.nih.gov) were “ulcerative colitis” (MeSH Terms) AND “human” [Organism] [12]. We select datasets with normal tissue samples and UC colon tissue samples: GSE87466, GSE75214, and GSE48958. The GSE87466 dataset was generated on the GPL13158 platform (HT_HG-U133_Plus_PM Affymetrix HT HG-U133+ PM Array Plate), and the GSE75214 and GSE48958 datasets were generated on the GPL6244 platform (HuGene-1_0-st Affymetrix Human Gene 1.0 ST Array transcript [gene] version). The GSE87466, GSE75214, and GSE48958 datasets contained 21, 11, and 8 normal tissue samples and 87, 97, and 13 UC colon tissue samples, respectively.
DATA PROCESSING AND SCREENING OF DEGS:
The MINiML formatted family files for each of the GEO datasets were downloaded. R language was used to extract the matrix file, and standardization was performed using quantiles. We used
COMMON DEGS ENRICHMENT ANALYSIS:
DEGs were subjected to Gene Ontology (GO) and Kyoto Encyclopedia of Gene and Genome (KEGG) enrichment analyses using the Database for Annotation, Visualization and Integrated Discovery (DAVID) 6.8 (
PPI NETWORK ANALYSIS:
PPI network analysis was performed using the Search Tool for the Retrieval of Interacting Genes (STRING) (
ENRICHMENT ANALYSIS OF MODULE GENES AND HUB GENES:
The Metascape Database (https://metascape.org/) [13] was used to perform enrichment analysis for genes in the 2 top-scoring modules. Metascape Database integrates multiple authoritative data resources including GO, KEGG, UniProt, and DrugBank, which can better complete pathway enrichment and annotation of biological processes. Reactome (https://reactome.org/) provides genetic pathway visualization, interpretation, and analysis. We used this pathway database for hub gene enrichment analysis. GeneCards (https://www.genecards.org/), which provides comprehensive biological information on annotated and predicted human genes, was used to search for single hub genes of CXCL8 and CXCL1.
Results
IDENTIFYING DEGS IN UC TISSUE AND NORMAL TISSUE:
We uniformly standardized the 3 datasets (Figure 1). We obtained 527, 293, and 125 DEGs from GSE87466, GSE75214, and GSE48958, respectively (Figure 2A). The GSE87466 dataset contained 347 upregulated DEGs and 180 downregulated DEGs. The GSE75214 and GSE48958 datasets contained 187 upregulated and 106 downregulated DEGs and 69 upregulated and 56 downregulated DEGs, respectively. The top 100 DEGs are displayed in heat maps (Figure 2B). All up- and downregulated DEGs were screened using Venn diagram software to identify common DEGs. A total of 52 upregulated and 38 downregulated common DEGs were obtained (Figure 2C).
GO BIOLOGICAL FUNCTIONS AND KEGG PATHWAY ENRICHMENT ANALYSIS OF COMMON DEGS:
The results of the GO enrichment analysis are shown in Figure 3. In the biological process analysis, DEGs were mainly involved in the inflammatory response, defense response, immune response, response to bacteria, and response to cytokine stimulus. In the cellular component analysis, DEGs were mainly enriched in extracellular regions and in the cell, membrane, and insoluble fractions. In the molecular function analysis, DEGs were mainly enriched in tetrapyrrole binding, cytokine activity, chemokine activity, heme binding, and calcium ion binding.
The main KEGG analysis results showed that DEGs were mainly concentrated in starch and sucrose metabolism and ATP-binding cassette (ABC) transporter pathways (Figure 3).
PPI NETWORK AND VISUALIZATION OF GENES:
We obtained a PPI network composed of 69 nodes and 180 edges (Figure 4A). Further analysis using the MCODE plugin identified 3 functional modules. The 2 top-scoring modules were CCL20 and CXCL8. The CCL20 module included CCL20, MMP1, CXCL13, C3, CXCL11, IDO1, NOS2, SOCS3, IL1RN, MMP3, MMP7, and TIMP1. They were closely connected with each other in the CCL20 module. CXCL8, CXCL1, IL-1β, PTGS2, LCN2, and CHI3L1 were more highly associated with another module (Figure 4B–4D). The hub genes identified using the cytoHubba plugin included CXCL8, CXCL1, IL-1β, PTGS2, TIMP1, CCL20, C3, MMP1, LCN2, and MMP3. Among them, CXCL8, CXCL1, and IL-1β were the top 3 hub genes (Figure 4E).
FURTHER ENRICHMENT ANALYSIS OF THE MODULE GENES AND HUB GENES:
CXCL8 and CCL20 module genes were further enriched using the Metascape database, which integrates GO function and KEGG pathway analyses (Figure 5A, 5B). CXCL8 module genes were mainly enriched in the cytokine-mediated signaling pathway, response to bacteria, regulation of inflammatory response, pregnancy (females), and positive regulation of response to external stimulus. CCL20 module genes were predominantly enriched in the cellular response to IL-1, positive regulation of angiogenesis, and antimicrobial humoral response. The KEGG pathway enrichment analysis showed that CXCL8 module genes were significantly enriched in IL-10 signaling, TNF signaling, and regulation of insulin-like growth factor. CCL20 module genes were mainly enriched in the IL-17, IL-4, and IL-13 signaling pathways.
The top 3 hub genes, CXCL8, CXCL1, and IL-1β, were subjected to further GO analysis (Table 1). In the biological process analysis, CXCL8 was mainly enriched in angiogenesis and antimicrobial humoral immune response mediated by antimicrobial peptides. CXCL1 was predominantly enriched in actin cytoskeleton organization and in the cellular response to lipopolysaccharide. IL-1β was significantly enriched in activation of MAPK activity and apoptotic processes. In the cellular component analysis, CXCL8, CXCL1, and IL-1β were mainly enriched in the extracellular region and in extracellular space. CXCL1 was also enriched in the specific granule lumen and tertiary granule lumen. IL-1β was also enriched in the cytosol, lysosome, and secretory granule. In the molecular function analysis, CXCL8 and CXCL1 were mainly enriched in chemokine activity, CXCR chemokine receptor binding, and IL-8 receptor binding. IL-1β was predominantly enriched in chemokine activity and integrin binding.
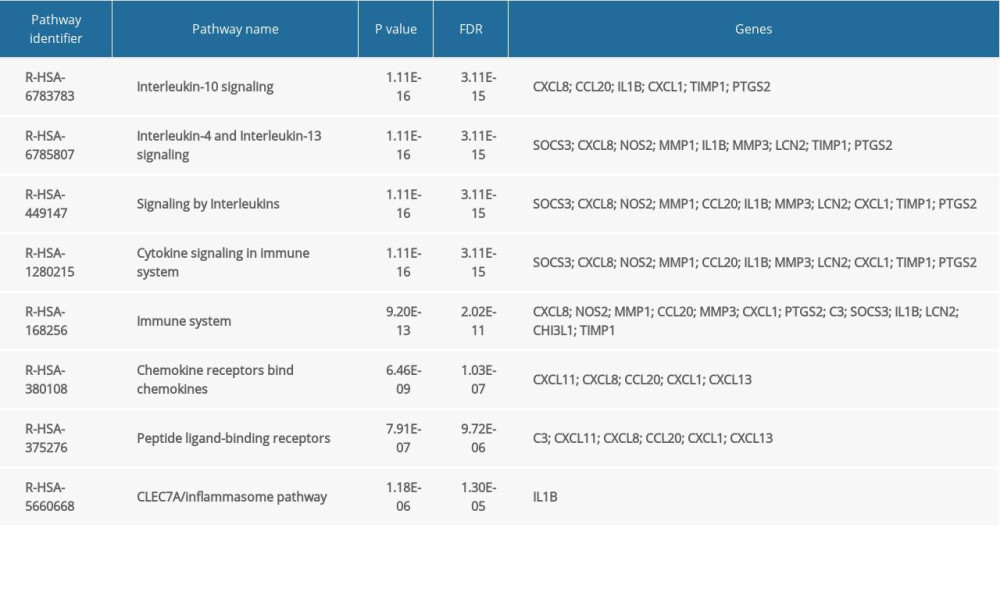
The main pathway enrichment results for hub genes are shown in Table 2. The top 3 hub genes were mainly enriched in IL-10, IL-4, and IL-13 signaling; signaling by interleukins; cytokine signaling in the immune system; and in the immune system. Among them, the P value of the IL-10 signaling pathway was the lowest (Figure 5C). This pathway is composed of 2 chemical components, 45 proteins, and 39 DNA and RNA molecules. The expression levels of 45 IL-10 signaling pathway genes in various tissues in the human body are shown in Figure 5D. CXCL8, CXCL1, and IL-1β are expressed in many organs including the colon, duodenum, appendix, and liver.
Discussion
This study screened gene UC and normal tissue expression datasets and identified 90 DEGs, including 3 functional modules and 1 hub gene module. Enrichment analysis found that the hub gene module was enriched in IL-10, IL-4, and IL-13 signaling pathways. In addition,
UC is a global, progressive, and complex disease with a long course and recurrent episodes. The incidence rates of UC are increasing worldwide, but the pathogenesis remains unclear and current treatments are not satisfactory. The relatively recent development of bioinformatics has led to the emergence of targeted molecular therapies for many diseases. Zhong et al. [14] used a bioinformatics analysis to show that the ATP-citrate lyase (ACLY) hub gene may be a molecular target in type 2 diabetes. Feng et al. [15] identified 4 significantly upregulated DEGs related to poor prognosis in ovarian cancer that could be potential therapeutic targets for patients with ovarian cancer. Similarly, exploring the molecular regulatory mechanism of UC occurrence and development by screening and analyzing key genes at the genetic level is expected to provide new directions for the precise prevention and treatment of UC in the clinic.
IL-10 is an anti-inflammatory cytokine that inhibits a broad spectrum of activated macrophage/monocyte functions. IL-10 plays an important role in UC. Previous studies [12,16] revealed that intestinal inflammation may be aggravated in individuals with IL-10 deficiency through inhibition of regulatory T cells/cytotoxic T-lymphocyte-associated protein 4 and through promotion of the IL-1β/T helper 2 cell pathway. Studies have shown that IL-10 can negatively regulate
Chemokine (C-X-C motif) ligand 8 (CXCL8), also known as IL-8 and neutrophil factor, can be secreted by lymphocytes, monocytes, and epithelial cells, and it plays an important role in the occurrence and development of tumors by promoting angiogenesis and immune cell infiltration [20]. Additionally, CXCL8 can recruit and activate neutrophils and granulocytes to migrate to the site of inflammation [21,22]. Heidarian et al. [23] found that changing the intestinal flora of patients with IBD can promote increased IL-8 expression and thereby increase the severity of the disease. Another study showed that IL-8 expression is increased in patients with IBD, especially in conjunction with UC [24]. These results are consistent with those presented here, which indicate that
CXCL1 is also known as NAP-3, CINC-1, and GRO-α. This protein was originally identified in melanoma, and it was later confirmed to be expressed in neutrophils, macrophages, and epithelial cells and to be involved in inflammation, angiogenesis, and wound-healing biological processes [25]. Alzoghaibi et al. [26] found that GRO-α levels were significantly higher in patients with IBD than in healthy controls, and that elevated GRO-α levels exacerbated IBD-related inflammation. This finding is consistent with the results presented by Mitsuyama et al. [27] showing that serum GRO-α is positively correlated with IBD activity and may be a marker for detecting IBD activity. In the present study,
Prostaglandin-endoperoxide synthase (PTGS), or cyclooxygenase, is a key enzyme in prostaglandin biosynthesis. It is involved in various biological processes including inflammation, reproduction, and tumor migration. Song et al. [31] showed that reducing
Chemokine (CC motif) ligand 20 (CCL20) is also known as human macrophage inflammatory protein 3α. The combination of CCL20 and CCR6 may be responsible for the immunosuppressive effect of marrow-derived suppressor cells [34]. Similarly, Woznicki et al. [35] found that the CCL20 and CCL5 chemokines are involved in the immune response in the human intestine. CCL20 may be an important regulatory molecule in the pathogenesis of IBD. Skovdahl et al. [36] found increased expression of CCL20 and CCR6 in colonic mucosal epithelial cells and peripheral blood mononuclear cells in patients with IBD. While our results indicate that CCL20 has less effect on UC occurrence and development than do the other 2 hub genes, it does not indicate that CCL20 is of less importance in UC. Confirmation of these results using a large number of samples and experiments is required.
There is overlap between UC and Crohn’s disease, which is why previous bioinformatics studies have focused on IBD. The current study used 3 GEO datasets that relied on the accurate identification of UC, which was not validated in this study. Larger datasets should be analyzed in future. This study was a bioinformatics study, and it did not validate the potential molecular biomarkers identified.
Conclusions
Bioinformatics analysis using 3 GEO datasets identified
Figures
 Figure 1. Standardization of gene expression. Standardization of gene expression in (A) GSE87466, (B) GSE75214, and (C) GSE48958 datasets. The blue bar represents the data before normalization, and the red bar represents the data after normalization.
Figure 1. Standardization of gene expression. Standardization of gene expression in (A) GSE87466, (B) GSE75214, and (C) GSE48958 datasets. The blue bar represents the data before normalization, and the red bar represents the data after normalization. 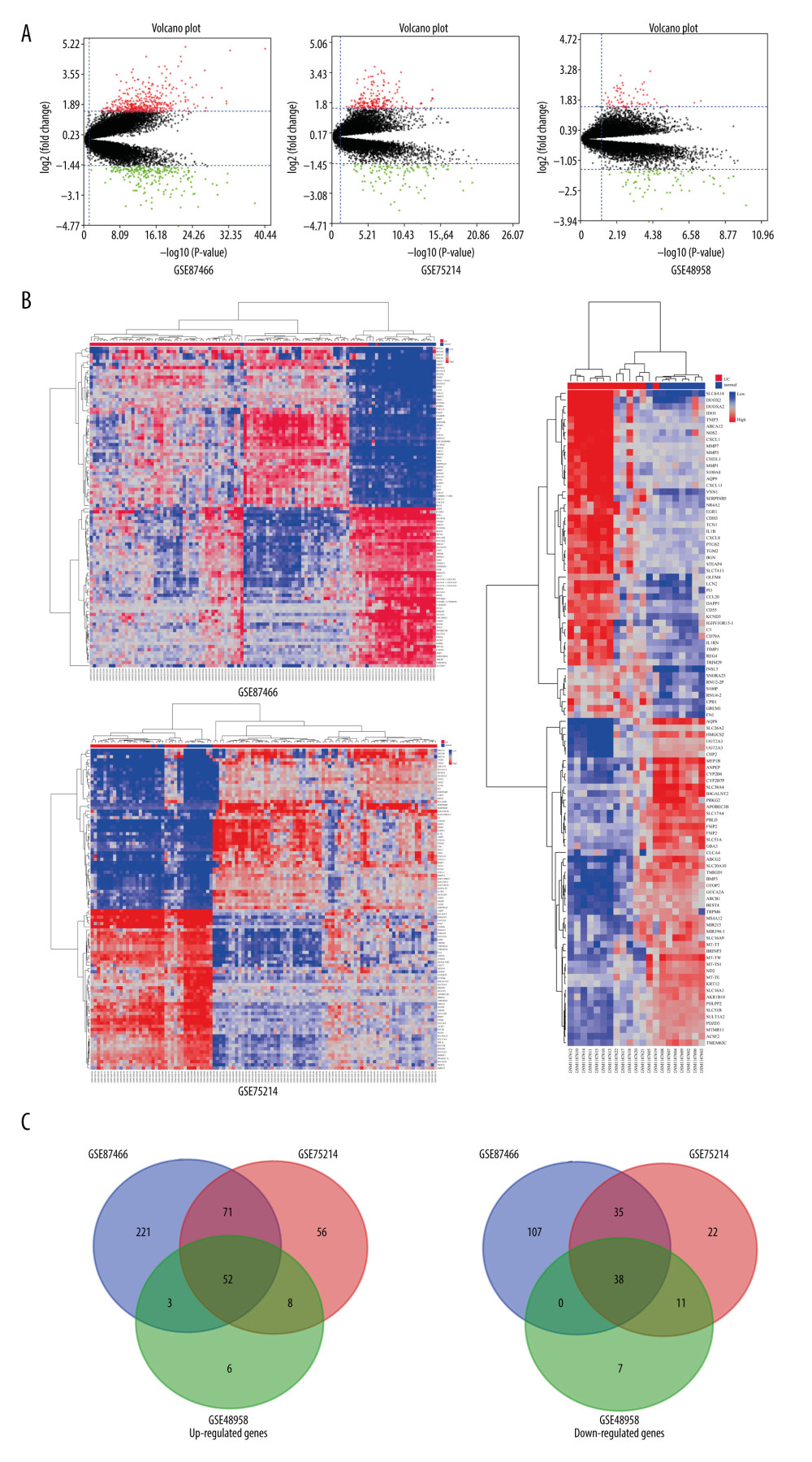 Figure 2. Screening results for the differentially expressed genes. (A) Volcano maps of gene expression in GSE87466, GSE75214, and GSE48958 datasets. Data points in red, green, and black represent genes with upregulated, downregulated, and not significantly different expression, respectively. (B) Heat maps of gene expression GSE87466, GSE75214, and GSE48958 datasets. (C) Venn diagram of common differentially expressed genes in GSE87466, GSE75214, and GSE48958 datasets.
Figure 2. Screening results for the differentially expressed genes. (A) Volcano maps of gene expression in GSE87466, GSE75214, and GSE48958 datasets. Data points in red, green, and black represent genes with upregulated, downregulated, and not significantly different expression, respectively. (B) Heat maps of gene expression GSE87466, GSE75214, and GSE48958 datasets. (C) Venn diagram of common differentially expressed genes in GSE87466, GSE75214, and GSE48958 datasets. 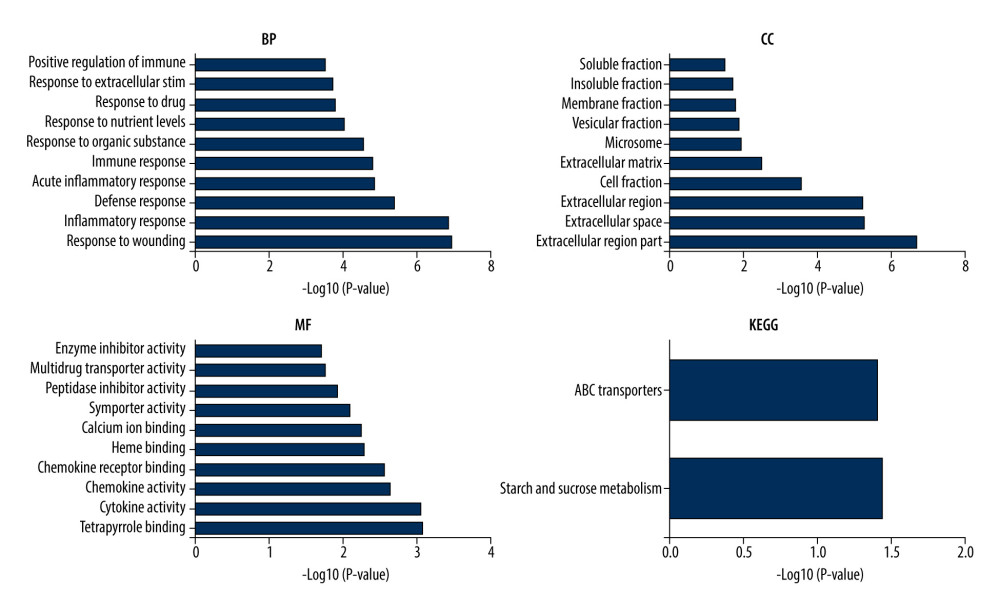 Figure 3. Gene ontology (GO) and Kyoto Encyclopedia of Gene and Genome (KEGG) pathway enrichment analysis for common differentially expressed genes.
Figure 3. Gene ontology (GO) and Kyoto Encyclopedia of Gene and Genome (KEGG) pathway enrichment analysis for common differentially expressed genes. 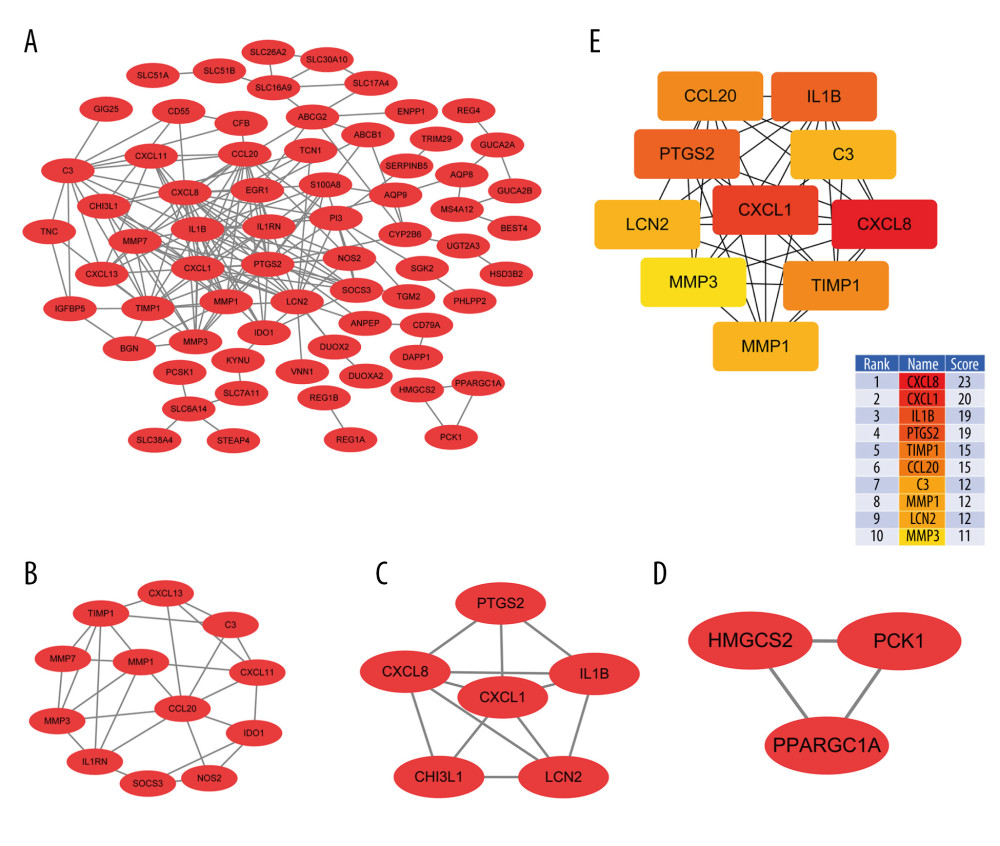 Figure 4. Protein–protein interaction (PPI) analysis. (A) PPI networks of the common differentially expressed genes. (B–D) The 3 top-scoring modules are shown from the top to bottom. (E) The PPI hub gene module.
Figure 4. Protein–protein interaction (PPI) analysis. (A) PPI networks of the common differentially expressed genes. (B–D) The 3 top-scoring modules are shown from the top to bottom. (E) The PPI hub gene module. 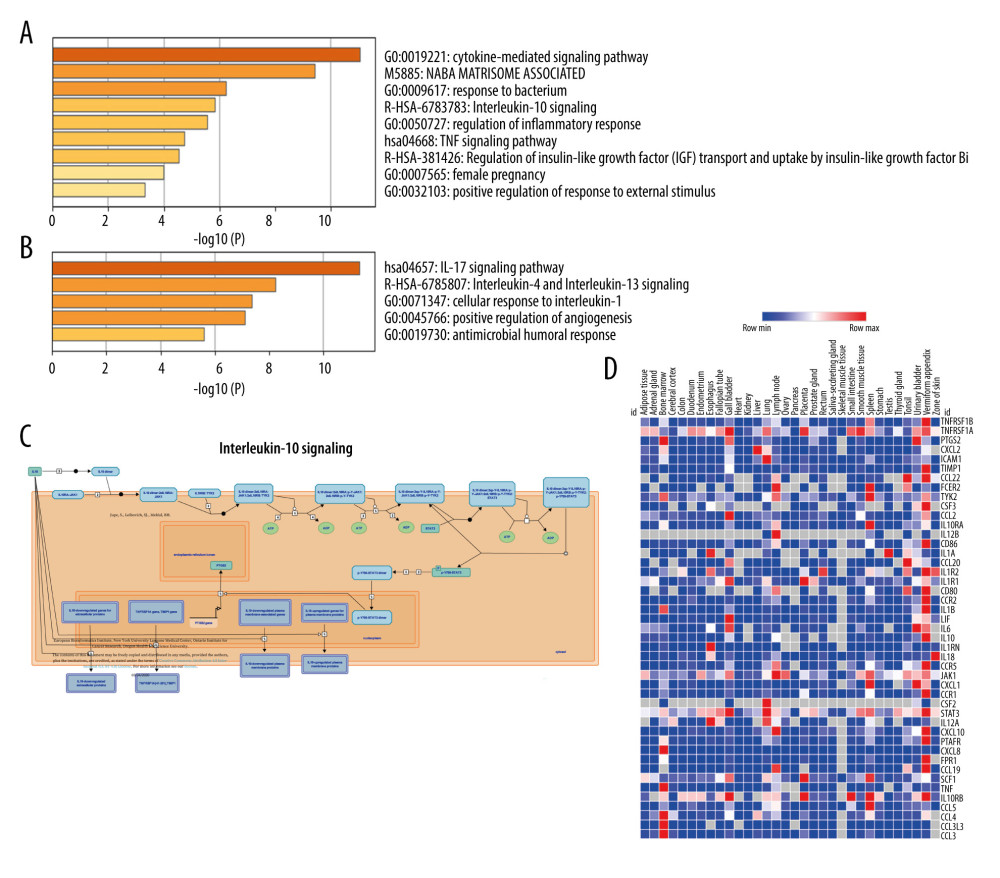 Figure 5. Metascape enrichment analysis of the top 2 modules and reactome pathway enrichment analysis for hub gene modules. (A) Metascape analysis of the CCL20 modules. (B) Metascape analysis of the CXCL8 modules. (C) Interleukin (IL)-10 signaling pathway. (D) Heat map of the expression of all proteins involved in the IL-10 signaling pathway in the human body.
Figure 5. Metascape enrichment analysis of the top 2 modules and reactome pathway enrichment analysis for hub gene modules. (A) Metascape analysis of the CCL20 modules. (B) Metascape analysis of the CXCL8 modules. (C) Interleukin (IL)-10 signaling pathway. (D) Heat map of the expression of all proteins involved in the IL-10 signaling pathway in the human body. 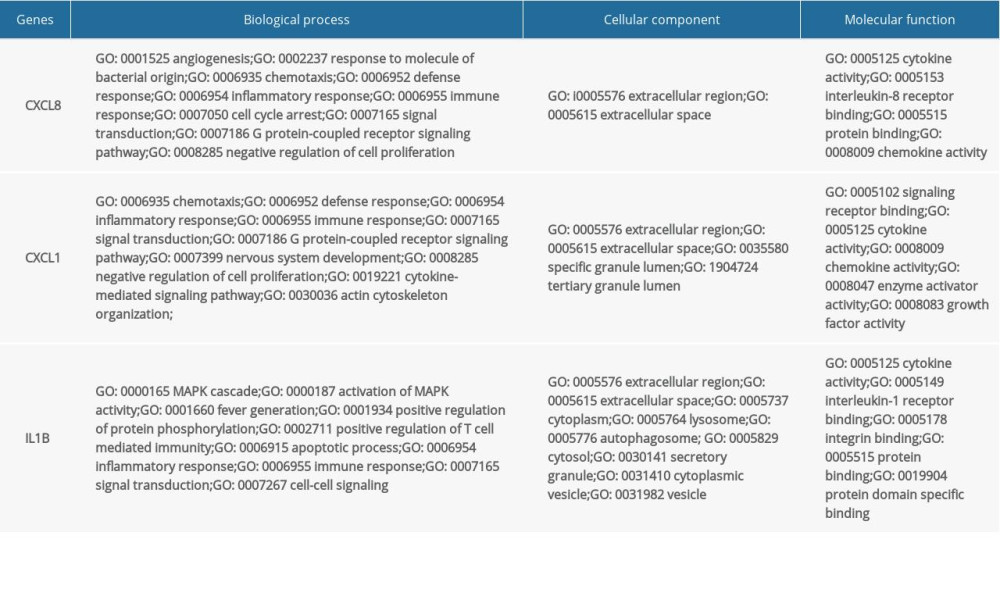
References
1. Ng SC, Tang W, Ching JY, Incidence and phenotype of inflammatory bowel disease based on results from the Asia-Pacific Crohn’s and colitis epidemiology study: Gastroenterology, 2013; 145; 158-65.e2
2. Xu M, Tao J, Yang Y, Ferroptosis involves in intestinal epithelial cell death in ulcerative colitis: Cell Death Dis, 2020; 11(2); 86
3. Ungaro R, Mehandru S, Allen PB, Ulcerative colitis: Lancet, 2017; 389; 1756-70
4. Guan Q, Zhang J, Recent advances: The imbalance of cytokines in the pathogenesis of inflammatory bowel disease: Mediators Inflamm, 2017; 2017 4810258
5. Kvorjak M, Ahmed Y, Miller ML, Cross-talk between colon cells and macrophages increases ST6GALNAC1 and MUC1-sTn expression in ulcerative colitis and colitis-associated colon cancer: Cancer Immunol Res, 2020; 8; 167-78
6. Eisenstein M, Ulcerative colitis: Towards remission: Nature, 2018; 563; S33
7. Mark-Christensen A, Erichsen R, Brandsborg S, Pouch failures following ileal pouch-anal anastomosis for ulcerative colitis: Colorectal Dis, 2018; 20; 44-52
8. Xu YY, He AQ, Liu G, Enteral nutrition combined with glutamine promotes recovery after ileal pouch-anal anastomosis in rats: World J Gastroenterol, 2018; 24; 583-92
9. Li KY, Wang JL, Xu YY, Intestinal barrier disruption in ileal pouchitis after ileal pouch-anal anastomosis in a rat model: Inflamm Bowel Dis, 2017; 23; 923-31
10. Cheng C, Hua J, Tan J, Identification of differentially expressed genes, associated functional terms pathways, and candidate diagnostic biomarkers in inflammatory bowel diseases by bioinformatics analysis: Exp Ther Med, 2019; 18; 278-88
11. Feng J, Gao Q, Liu Q, Integrated strategy of differentially expressed genes associated with ulcerative colitis: Mol Med Rep, 2017; 16; 7479-89
12. Osadnik T, Strzelczyk JK, Lekston A, The association of functional polymorphisms in genes encoding growth factors for endothelial cells and smooth muscle cells with the severity of coronary artery disease: BMC Cardiovasc Disord, 2016; 16; 218
13. Zhou Y, Zhou B, Pache L, Metascape provides a biologist-oriented resource for the analysis of systems-level datasets: Nat Commun, 2019; 10; 1523
14. Zhong M, Wu Y, Ou W, Identification of key genes involved in type 2 diabetic islet dysfunction: A bioinformatics study: Biosci Rep, 2019; 39(5); BSR20182172
15. Feng H, Gu ZY, Li Q, Identification of significant genes with poor prognosis in ovarian cancer via bioinformatical analysis: J Ovarian Res, 2019; 12; 35
16. Ye M, Joosse ME, Liu L, Deletion of IL-6 exacerbates colitis and induces systemic inflammation in IL-10-deficient mice: J Crohns Colitis, 2019; 14(6); 831-40
17. Kopydlowski KM, Salkowski CA, Cody MJ: J Immunol, 1999; 163; 1537-44
18. de Waal MR, Figdor CG, Huijbens R, Effects of IL-13 on phenotype, cytokine production, and cytotoxic function of human monocytes. Comparison with IL-4 and modulation by IFN-gamma or IL-10: J Immunol, 1993; 151; 6370-81
19. Niiro H, Otsuka T, Kuga S, IL-10 inhibits prostaglandin E2 production by lipopolysaccharide-stimulated monocytes: Int Immunol, 1994; 6; 661-64
20. Stanisavljevic L, Assmus J, Storli KE, CXCR4, CXCL12 and the relative CXCL12-CXCR4 expression as prognostic factors in colon cancer: Tumour Biol, 2016; 37; 7441-52
21. Liu Q, Yu S, Li A, Targeting interlukin-6 to relieve immunosuppression in tumor microenvironment: Tumour Biol, 2017; 39(6) 1010428317712445
22. Wu K, Yu S, Liu Q, The clinical significance of CXCL5 in non-small cell lung cancer: Onco Targets Ther, 2017; 10; 5561-73
23. Heidarian F, Alebouyeh M, Shahrokh S, Altered fecal bacterial composition correlates with disease activity in inflammatory bowel disease and the extent of IL8 induction: Curr Res Transl Med, 2019; 67; 41-50
24. Neubauer K, Matusiewicz M, Bednarz-Misa I, Diagnostic potential of systemic eosinophil-associated cytokines and growth factors in IBD: Gastroenterol Res Pract, 2018; 2018 7265812
25. Yu S, Yi M, Xu L, CXCL1 as an unfavorable prognosis factor negatively regulated by DACH1 in non-small cell lung cancer: Front Oncol, 2019; 9; 1515
26. Alzoghaibi MA, Al-Mofleh IA, Al-Jebreen AM, Neutrophil chemokines GCP-2 and GRO-alpha in patients with inflammatory bowel disease: J Dig Dis, 2008; 9; 144-48
27. Mitsuyama K, Tsuruta O, Tomiyasu N, Increased circulating concentrations of growth-related oncogene (GRO)-alpha in patients with inflammatory bowel disease: Dig Dis Sci, 2006; 51; 173-77
28. Tian T, Lofftus S, Pan Y, IL1alpha antagonizes IL1beta and promotes adaptive immune rejection of malignant tumors: Cancer Immunol Res, 2020; 8(5); 660-71
29. Zhong XS, Winston JH, Luo X, Neonatal colonic inflammation epigenetically aggravates epithelial inflammatory responses to injury in adult life: Cell Mol Gastroenterol Hepatol, 2018; 6; 65-78
30. De Santis S, Kunde D, Galleggiante V, TNFalpha deficiency results in increased IL-1beta in an early onset of spontaneous murine colitis: Cell Death Dis, 2017; 8; e2993
31. Song JL, Lee JS, Kim HY, Dietary mixed cereal grains ameliorate the azoxymethane and dextran sodium sulfate-induced colonic carcinogenesis in C57BL/6J mice: J Med Food, 2020; 23(4); 440-52
32. Venè R, Costa D, Augugliaro R, Evaluation of glycosylated PTGS2 in colorectal cancer for NSAIDS-based adjuvant therapy: Cells, 2020; 9(3); 683
33. Grabauskas G, Wu X, Gao J, Prostaglandin E2, produced by mast cells in colon tissues from patients with irritable bowel syndrome, contributes to visceral hypersensitivity in mice: Gastroenterology, 2020; 158(8); 2195-207.e6
34. Zhao Y, Lu F, Ye J: Front Immunol, 2019; 10; 3104
35. Woznicki JA, Flood P, Bustamante-Garrido M, Human BCL-G regulates secretion of inflammatory chemokines but is dispensable for induction of apoptosis by IFN-gamma and TNF-alpha in intestinal epithelial cells: Cell Death Dis, 2020; 11(1); 68
36. Skovdahl HK, Damås JK, Granlund AVB, C-C motif ligand 20 (CCL20) and C-C motif chemokine receptor 6 (CCR6) in human peripheral blood mononuclear cells: dysregulated in ulcerative colitis and a potential role for CCL20 in IL-1β release: Int J Mol Sci, 2018; 19(10); 3257
Figures
Figure 1. Standardization of gene expression. Standardization of gene expression in (A) GSE87466, (B) GSE75214, and (C) GSE48958 datasets. The blue bar represents the data before normalization, and the red bar represents the data after normalization.Figure 2. Screening results for the differentially expressed genes. (A) Volcano maps of gene expression in GSE87466, GSE75214, and GSE48958 datasets. Data points in red, green, and black represent genes with upregulated, downregulated, and not significantly different expression, respectively. (B) Heat maps of gene expression GSE87466, GSE75214, and GSE48958 datasets. (C) Venn diagram of common differentially expressed genes in GSE87466, GSE75214, and GSE48958 datasets.Figure 3. Gene ontology (GO) and Kyoto Encyclopedia of Gene and Genome (KEGG) pathway enrichment analysis for common differentially expressed genes.Figure 4. Protein–protein interaction (PPI) analysis. (A) PPI networks of the common differentially expressed genes. (B–D) The 3 top-scoring modules are shown from the top to bottom. (E) The PPI hub gene module.Figure 5. Metascape enrichment analysis of the top 2 modules and reactome pathway enrichment analysis for hub gene modules. (A) Metascape analysis of the CCL20 modules. (B) Metascape analysis of the CXCL8 modules. (C) Interleukin (IL)-10 signaling pathway. (D) Heat map of the expression of all proteins involved in the IL-10 signaling pathway in the human body. Tables
In Press
12 Mar 2024 : Clinical Research
Metabolomic Alterations in Methotrexate Treatment of Moderate-to-Severe PsoriasisMed Sci Monit In Press; DOI: 10.12659/MSM.943360
14 Mar 2024 : Clinical Research
Renal Dysfunction Increases Risk of Adverse Cardiovascular Events in 5-Year Follow-Up Study of Intermediate...Med Sci Monit In Press; DOI: 10.12659/MSM.943956
15 Mar 2024 : Clinical Research
Impact of One-Lung Ventilation on Oxygenation and Ventilation Time in Thoracoscopic Heart Surgery: A Compar...Med Sci Monit In Press; DOI: 10.12659/MSM.943089
14 Mar 2024 : Clinical Research
Differential DHA and EPA Levels in Women with Preterm and Term Births: A Tertiary Hospital Study in IndonesiaMed Sci Monit In Press; DOI: 10.12659/MSM.943895
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952