26 March 2021: Animal Study
Salvianic Acid A Regulates High-Glucose-Treated Endothelial Progenitor Cell Dysfunction via the AKT/Endothelial Nitric Oxide Synthase (eNOS) Pathway
Yanhua Guan12ABCDEF, Xu Wang13ABCDEF*DOI: 10.12659/MSM.928153
Med Sci Monit 2021; 27:e928153






Abstract
BACKGROUND: The primary cause of death in patients with diabetes mellitus (DM) is diabetic macroangiopathy, a complication that related to the function and number of endothelial progenitor cells (EPCs). Salvianic acid A (SAA) is a water-soluble active ingredient of Salvia miltiorrhiza, a traditional Chinese medicine used to treat cardiovascular diseases. The purpose of this study was to explore the effects of SAA on the function of rat EPCs cultured in vitro in a high-glucose environment.
MATERIAL AND METHODS: Bone marrow-derived EPCs from 40 Sprague-Dawley rats were identified by fluorescence staining. Cell viability, apoptosis, tube formation, lactated dehydrogenase (LDH) release, and nitric oxide (NO) production were detected by 3-[4,5-dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide assay, flow cytometry, tube formation, LDH, and 3-amino,4-aminomethyl-2’,7’-difluorescein, and diacetate assays, respectively. The expression levels of proteins were examined by western blotting.
RESULTS: Cultured EPCs showed a cobblestone morphology and positive expression of Dil-ac-LDL and FITC-UEA-1. High glucose impaired cell viability. Different concentrations of SAA had no significant effect on EPC viability. SAA reduced the apoptosis rate and LDH release, but promoted tube formation, viability, and NO production in high-glucose-treated EPCs. The ratios of p-AKT/AKT and p-eNOS/eNOS in high-glucose-treated EPCs were elevated by SAA. Phosphoinositide 3-kinase inhibitor LY294002 blocked the rescue effects of SAA on high-glucose-treated EPCs.
CONCLUSIONS: SAA protected EPCs against high-glucose-induced dysfunction via the AKT/eNOS pathway.
Keywords: Diabetes Complications, endothelial cells, Nitric Oxide Synthase, Diabetes Mellitus, endothelial progenitor cells, Glucose, Lactates, Nitric Oxide Synthase Type III, Proto-Oncogene Proteins c-akt, Salvia
Background
Diabetes mellitus (DM) is a metabolic disorder [1]. The incidence of DM has shown a significantly growing trend worldwide, with the number of patients with DM at about 463 million in 2019 [2]. DM is accompanied by multiple complications such as macroangiopathy, retinopathy, cardiomyopathy and renal failure, with macroangiopathy characterized by atherosclerosis, with thrombosis being its main complication [3]. Hyperglycemia in the presence of DM impairs the endothelium, causes endothelial cell apoptosis, and damages the integrity of the vascular endothelium [4] and are considered to be the initiating factors of macroangiopathy in DM [4,5]. The above complications are related, to a greater or lesser degree, to the reduction of endothelial progenitor cells (EPCs) [6].
EPCs have generated considerable research interest owing to their extensive therapeutic applications in cell therapy [7]. In one study, vascular endothelial cells were repaired by adjacent endothelial cells after continuous stimulus damage. However, to heal the local irreparable damage, EPCs must first mobilize from the bone marrow to the peripheral circulation, then to the lesion, where they differentiate into vascular endothelial cells [8]. Studies have reported that EPCs contribute to thrombus resolution and recanalization and repair the blood vessel damage caused by diabetes [8,9]. Although EPCs possess great potential in cell therapy, their functions can be impeded by cardiovascular diseases, dilation diseases, and transplanted vascular diseases [10]. In addition, high glucose (HG) destroys the biological function of EPCs. One study reported that HG increased the accumulation of reactive oxygen species (ROS) in the body, which could not be removed by cells, resulting in oxidative stress damage and impaired metabolism of EPCs [11]. Thus, it is critical to explore a novel mechanism to improve the functions of EPCs.
Salvianic acid A (SAA), also known as 3-(3,4-dihydroxyphenyl)-2-hydroxypropanoic acid, is an aromatic acid compound extracted from the water-soluble ingredients of danshen, the dried root of
Nitric oxide (NO) is essential for maintaining normal vascular tone [17]. In addition, NO is a free foundation that can diffuse through cell membranes and, together with certain nitrogen-containing derivatives, participates in bodily functioning through interactions with biological information molecules [18]. Many studies have confirmed that NO not only participates in the regulation of vascular tension, but also inhibits platelet aggregation, monocyte adhesion, inflammation, and oxidation [19]. In this study, we also explored the effect of SAA on HG-treated EPCs by regulating NO production.
Material and Methods
ANIMALS AND REAGENTS:
All animal experiments were conducted with the approval of the Animal Ethics Committee of the Nanjing Hospital of Chinese Medicine, affiliated with the Nanjing University of Chinese Medicine (approval No: DS201911035). Sprague-Dawley (SD) rats were purchased from Guangdong Medical Laboratory Animal Center (
The SD rats were euthanized by cervical dislocation after anesthesia with 1% pentobarbital (40 mg/kg) (76-74-4, Merck, Germany), and then the bone marrow cavity was washed with phosphate buffered saline (PBS) (C0221A, Beyotime, China) until the washing solution became colorless and transparent. The lavage fluid from the bone marrow cavity was collected in a 15 mL centrifuge tube (430791, Corning, USA) and centrifuged. The supernatant and fat were removed, and cells were resuspended with 5 mL of PBS. An amount of 5 mL of lymphocyte separation solution (P6030, Solarbio, China) and the prepared cell suspension were added to a new 15 mL centrifuge tube. Mononuclear cells were obtained by density gradient centrifugation and seeded into 6-well plates, which were coated with fibronectin (54008, Coring, USA). The cells were grown in M199 medium (11150067, Gibco, USA), which contained 20% fetal bovine serum (10091, Gibco, USA), 20 ng/mL vascular endothelial growth factor (VEGF) (PRG0113, Thermo Fisher, USA), 5 ng/mL basic fibroblast growth factor (P6348, Beyotime, China), and 1% penicillin-streptomycin (15140-122, Gibco, USA).
IDENTIFICATION OF EPCS:
An optical microscope at 100× magnification (BX53M, Olympus, Japan) was used to observe the morphological changes of cells after 4 days, 8 days, and 14 days of cell culture. EPCs were allowed to differentiate and grow in vitro for 2 weeks to reach 70% fusion. Next, 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein (Dil-ac-LDL) (H7970, Solarbio, China) was diluted with medium to 30 μg/mL, and the dilution was added to the cells and incubated at 37 °C for 4 h in an incubator (Forma Steri-Cycle, Thermo Scientific, USA). After washing twice with PBS, the cells were fixed with 4% paraformaldehyde (P0099, Beyotime, China) for 15 min. Then, 400 μL of 10 μg/mL fluorescein isothiocyanate-Ulex europaeus agglutinin-1 (FITC-UEA-1) (L9006, Merck, Germany) was added to the specimens and incubated at 37°C for 1 h in an incubator. A fluorescence microscope at 200× magnification (IX73, Olympus, Japan) was used to observe the staining results.
CELL CIABILITY ASSAY:
A 3-[4,5-dimethylthylthiazol-2-yl]-2, 5 diphenyltetrazolium bromide (MTT) assay kit (ab211091, Abcam, UK) was used to detect cell viability. For detection of the effects of HG on the EPCs, the cells were divided into 2 groups, according to previous research reports [17, 18]: a control group (5.5 mmol/L glucose and 25 mmol/L mannitol) and an HG group (30 mmol/L glucose). The glucose (921-60-8, Chemsynlab, China) and mannitol (M8140, Solarbio, China) were purchased from Beijing Chemsynlab Co., Ltd. (http://www.chemsynlab.com/) and Beijing Solarbio Science & Technology Co., Ltd. (http://www.solarbio.com/), respectively. An amount of 5×103 EPCs were seeded into each well of 96-well plates and incubated for 1 day, 3 days, and 7 days. For detection of the effects of SAA on EPCs, cells were treated with different concentrations of SAA (0, 1, 5, 10, 25, 50, and 100 μmol/L) and incubated for 24 h. To detect the effects of SAA on HG-treated EPCs, the cells were divided into 4 groups: a control group, HG group, HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h), and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h). To detect the effects of LY294002 on the HG-treated EPCs, the cells were divided into 4 groups: a control group, HG group, HG+SAA group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured by 30 mmol/L glucose for 24 h), and HG+SAA+LY294002 group (cells were pretreated with 100 μmol/L SAA and LY294002 for 24 h and 1 h, respectively, and then cultured by 30 mmol/L glucose for 24 h). After incubation, 50 μL serum-free medium and 50 μL MTT reagent were added to each well. The 96-well plates were placed in an incubator for 3 h. Subsequently, the MTT reagent-medium mixture was removed and 150 μL of MTT solvent was added into each well, followed by shaking for 15 min on an orbital shaker (Solaris 4000 Shaker, Thermo Fisher, USA). The absorbance was examined by a multi-detection reader (SpectraMax5, Molecular Devices, USA) at 590 nm.
CELL APOPTOSIS ASSAY:
Apoptosis was detected by an Annexin V-FITC apoptosis detection kit (C1062S, Beyotime, China). An amount of 5×105 cells and 195 μL of Annexin V-FITC binding solution was added into a centrifuge tube. Subsequently, 10 μL of Annexin V-FITC and 5 μL of propidium iodide were added to the centrifuge tube and mixed gently, followed by incubation at room temperature in the dark for 20 min. Then, the sample was detected by a flow cytometer (DxFLEX, Beckman, USA) and data were analyzed by CytExpert software (Beckman, USA).
TUBE FORMATION ASSAY:
Pre-cooled matrigel (354230, Corning, USA) was mixed with serum-free medium, and the mixture was then used to coat 24-well plates. The plates were placed into a 37 °C incubator for 30 min to solidify the matrigel. Treated cells were dispensed into the plates at 2×104 per well and incubated for 24 h. An inverted microscope (BX53M, Olympus, Japan) was used to take photographs.
LACTATED DEHYDROGENASE ACTIVITY ASSAY:
A lactated dehydrogenase (LDH) assay kit (C0017, Beyotime, China) was used to detect cytotoxicity. An amount of 3×103 cells were inoculated in a 96-well plate until the cell density reached 80%. After the cells were treated according to the experimental design, the supernatant was removed. Next, 150 μL of LDH release reagent was added to the culture plate and incubated for 1 h in an incubator. The supernatant was collected and mixed with 60 μL of LDH testing reagent. After incubation in the dark for 30 min at room temperature, the absorbance was measured at 490 nm by a multi-detection reader.
NO PRODUCTION:
To detect the production of NO, 3-Amino,4-aminomethyl-2′,7′-difluorescein, diacetate (DAF-FM DA) kits (S0019, Beyotime, China) were used. DAF-FM DA was diluted with the DAF-FM DA dilution provided with the kits at a ratio of 1: 1000 to a final concentration of 5 μmol/L. Cells were collected and resuspended with the diluted DAF-FM DA to a concentration of 1×107/mL. Then, the cells were incubated for 20 min at 37°C. The fluorescence intensity at an excitation wavelength of 495 nm and an emission wavelength of 515 nm was detected using a multi-detection reader.
WESTERN BLOTTING:
Western blotting was performed as previously described [20]. Proteins were collected using RIPA buffer (P0013B, Beyotime, China), protease inhibitors (p1005, Beyotime, China), and phenylmethylsulfonyl fluoride (ST505, Beyotime, China), quantified by a protein quantification kit (P0011, Beyotime, China), and separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Then, electrophoresis was conducted to transfer the proteins into a PVDF membrane (160-0184, Bio-Rad, USA). The membrane was blocked by 5% dried skimmed milk powder for 1 h and then incubated with antibodies against Akt (1: 500, 56kDa, ab8805, Abcam, UK), phosphorylated (p)-Akt (1: 1000, 56kDa, ab38449, Abcam, UK), endothelial nitric oxide synthase (eNOS) (1: 1000, 133kDa, ab76198, Abcam, UK), p-eNOS (1: 500, 140kDa, ab76199, Abcam, UK), and GAPDH (1: 1000, 36kDa, ab8245, Abcam, UK). After incubation for 24 h, the membrane was washed by 1% Tris-buffered saline with Tween 20 and incubated with goat anti-rabbit secondary antibody (1: 10 000, ab205718, Abcam, UK) or goat anti-mouse secondary antibody (1: 10000, ab6789, Abcam, UK). An ECL luminescence kit (PE0010, Solarbio, China) and a gel imaging system (FluorChem FC3, Alpha, USA) were used to expose the membrane and visualize the protein bands, respectively. ImageJ2x (Rawak Software, Germany) was used to analyze the results.
STATISTICAL ANALYSES:
Data were analyzed by Graph Prism v8.0 (Graphpad Software, California, USA) and SPSS version 20.0 (SPSS, Chicago, USA). Comparisons between groups were analyzed by one-way ANOVA and t test (the experiment shown in Figure 1C). All results were shown as mean±standard. The level of significance was defined as P<0.05.
Results
IDENTIFICATION OF BONE MARROW-DERIVED EPCS:
EPCs showed a round or elliptical shape after cultivation for 4 days, exhibited a fusiform or round shape after 8 days of culture, and eventually presented a cobblestone morphology after culture for 2 weeks (Figure 1A). Cells with positive expression of Dil-ac-LDL and FITC-UEA-1 were identified as differentiated EPCs (Figure 1B). To detect the effects of HG and SAA on cell viability, MTT assay was conducted. EPC viability was strongly suppressed in the HG group (Figure 1C, P<0.01), whereas different concentrations of SAA had no significant effect on EPC viability (about 100%, Table 1, P>0.05). Therefore, we chose 50 μmol/L and 100 μmol/L SAA for subsequent experiments.
SAA AT DIFFERENT CONCENTRATIONS ALLEVIATED THE EFFECTS OF HG ON APOPTOSIS, TUBE FORMATION, VIABILITY, LDH, AND NO RELEASE IN EPCS:
HG greatly elevated the cell apoptosis rate (20%); however, 50 μmol/L (15%) and 100 μmol/L (8%) of SAA reversed the elevation of the rate of cell apoptosis by HG (Figure 2A, 2B, P<0.05). In addition, the inhibitory effect of 100 μmol/L of SAA on the cell apoptosis rate in an HG environment was stronger than that of 50 μmol/L of SAA. Similarly, the branch points, cell viability, and NO production were inhibited and the level of LDH (150%) was promoted in the HG group; however, SAA reversed these effects in a dose-dependent manner (Figure 2C–2G, P<0.05).
EFFECTS OF SAA AT DIFFERENT CONCENTRATIONS ON THE EXPRESSION LEVELS OF AKT/ENOS PATHWAY-RELATED PROTEINS IN THE HG ENVIRONMENT:
To investigate the underlying mechanism of the effect of SAA on the Akt/eNOS pathway, western blot analysis was performed to evaluate the expression levels of Akt/eNOS pathway-related proteins. We found that the expressions of p-Akt (0.3) and p-eNOS (0.2) were inhibited by HG, whereas this inhibition was blocked by SAA in a dose-dependent manner (Figure 3A, 3B, P<0.01). However, there was no significant difference in the expressions of Akt and eNOS between the groups. Meanwhile, the ratios of p-Akt/Akt and p-eNOS/eNOS in the HG group (0.4 and 0.3, respectively) were more than 2 times lower than those in the control group (1.2 and 0.5, respectively). The inhibitory effects of HG on the ratios of p-Akt/Akt and p-eNOS/eNOS were reversed by SAA at 50 μmol/L (0.6 and 0.6, respectively) and 100 μmol/L (1.1 and 0.9, respectively) (Figure 3C, 3D, P<0.05).
EFFECTS OF COMBINED SAA AND PI3K INHIBITOR ON THE EXPRESSIONS OF AKT/ENOS PATHWAY-RELATED PROTEINS IN THE HG ENVIRONMENT:
Similar to the previous results, HG reduced the expressions of p-AKT and p-eNOS (0.3 and 0.25), SAA reversed the inhibitory effect of HG (0.9 and 0.8), and PI3K inhibitor LY294002 eliminated the effect of SAA (Figure 4A, 4B, P<0.001). Analogously, after using PI3K inhibitors, the effect of SAA on restoring the ratios of p-Akt/Akt and p-eNOS/eNOS affected by HG was abolished (Figure 4C, 4D, P<0.001).
EFFECTS OF COMBINED SAA AND PI3K INHIBITOR ON VIABILITY AND LDH AND NO PRODUCTION IN HG-TREATED EPCS:
HG alone inhibited cell viability and NO production and increased the LDH level. SAA changed the regulation of HG on cells, leading to increased cell viability and NO production and a reduced LDH level. However, the combination of PI3K inhibitor and SAA once again decreased cell viability and NO production and elevated LDH levels (Figure 4E, 4G, P<0.05).
Discussion
The main cause of death and disability in DM is chronic complications, especially diabetic macroangiopathy, which is a huge burden for patients [21]. EPC dysfunction is a possible cause of diabetes-induced macroangiopathy because of its participation in endothelial dysfunction and angiogenesis [6]. Therefore, improving EPC dysfunction has become a novel and hot research topic. Previous research has reported that metformin, insulin, and other commonly used drugs for diabetes may increase the number of EPCs to regulate EPC dysfunction [22]. In our present study, we first demonstrated the effect of SAA on improving EPC dysfunction in HG treatment. The EPCs of rats are generally derived from the peripheral blood, bone marrow, and spleen. Because the largest number of EPCs are in the bone marrow, we isolated and cultured the stem cells from the femur and tibia bone marrow cavities. After culturing the cells for 2 weeks, we observed that the cells showed a cobblestone-like morphology, which was consistent with the results of previous experiments [23]. Next, EPCs were identified by Dil-ac-LDL and FITC-UEA-1 dual fluorescence staining.
Previous studies have shown that NO promotes the mobilization of EPCs, whereas HG may cause the reduction of NO production to reduce proliferation and increase apoptosis in endothelial cells, eventually leading to macroangiopathy in DM patients [24,25]. SAA, a water-soluble active ingredient extracted from
The enzyme eNOS has a key role in regulating NO production and is an important target for the Akt/eNOS signaling pathway [31,32]. Studies have shown that HG greatly alleviated the phosphorylation level of eNOS and the bioavailability of NO [24]. VEGF activated eNOS through the AKT-dependent eNOS phosphorylation pathway, thereby increasing the number of circulating endothelial cells [33]. In addition, the activation of the Akt/eNOS pathway promoted the proliferation, metastasis, and vascular repairing ability of EPCs [34]. Crocetin elevated the bioavailability of NO through the Akt/eNOS pathway to restore the function of HG-treated EPCs [35]. Therefore, we determined the expression levels of Akt/eNOS pathway-related proteins. Our results indicated that SAA restored the ratios of p-Akt/Akt and p-eNOS/eNOS reduced by HG. Thus, we speculated that SAA may play a regulatory role in HG-treated EPCs through the Akt/eNOS pathway. To verify this hypothesis, we used the PI3K inhibitor LY294002 to inhibit the PI3K/AKT signaling pathway. In line with our expectations, LY294002 hindered the effect of SAA on restoring the functions of HG-treated EPCs in the expressions of Akt/eNOS pathway-related proteins, cell viability, and LDH and NO production, suggesting that SAA mitigated the regulatory effect of HG on EPCs via the Akt/eNOS pathway.
Conclusions
In conclusion, our study showed that SAA improved the dysfunction of EPCs in an HG environment by restoring the AKT-eNOS pathway, which may provide promising strategies for the treatment of DM and its complications. Nonetheless, studies increasingly indicate that DM is associated with a proinflammatory status, and its inflammation action mechanism suppresses EPC proliferation [36,37]. Also, SAA has been found to display anti-inflammatory pharmacological activity [38]. Hence, we will perform further experiments to explore the anti-inflammatory function of SAA in HG-treated EPCs. Studies have shown that endothelial cell dysfunction can be traced back to the mitochondrial oxidative stress caused by high levels of glucose and changes in the RhoA/ROCK/Akt signaling pathway [39]. However, the present study is a preliminary one, and we will continue to explore the influence of SAA on the EPC function of cultured rats by influencing other signaling pathways and improving the separation and culture of EPCs.
Figures
![Identification of bone marrow-derived endothelial progenitor cells (EPCs) and effects of high glucose (HG) or different concentrations of salvianic acid A (SAA) on cell viability. (A) Morphology of EPCs derived from bone marrow. Scale bar=100 μm. (B) 1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein (Dil-ac-LDL) and fluorescein isothiocyanate-Ulex europaeus agglutinin-1 (FITC-UEA-1) double staining for identification of EPCs. Scale bar=50 μm. (C) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay was performed to determine the effects of HG (30 mmol/L) on EPC viability at 1 day, 3 days and 7 days after culture. (Forty rats were used in this experiment.) ** P<0.01 compared to the control group.](https://jours.isi-science.com/imageXml.php?i=medscimonit-27-e928153-g001.jpg&idArt=928153&w=1000) Figure 1. Identification of bone marrow-derived endothelial progenitor cells (EPCs) and effects of high glucose (HG) or different concentrations of salvianic acid A (SAA) on cell viability. (A) Morphology of EPCs derived from bone marrow. Scale bar=100 μm. (B) 1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein (Dil-ac-LDL) and fluorescein isothiocyanate-Ulex europaeus agglutinin-1 (FITC-UEA-1) double staining for identification of EPCs. Scale bar=50 μm. (C) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay was performed to determine the effects of HG (30 mmol/L) on EPC viability at 1 day, 3 days and 7 days after culture. (Forty rats were used in this experiment.) ** P<0.01 compared to the control group.
Figure 1. Identification of bone marrow-derived endothelial progenitor cells (EPCs) and effects of high glucose (HG) or different concentrations of salvianic acid A (SAA) on cell viability. (A) Morphology of EPCs derived from bone marrow. Scale bar=100 μm. (B) 1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein (Dil-ac-LDL) and fluorescein isothiocyanate-Ulex europaeus agglutinin-1 (FITC-UEA-1) double staining for identification of EPCs. Scale bar=50 μm. (C) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay was performed to determine the effects of HG (30 mmol/L) on EPC viability at 1 day, 3 days and 7 days after culture. (Forty rats were used in this experiment.) ** P<0.01 compared to the control group. ![Salvianic acid A (SAA) at different concentrations regulated apoptosis, tube formation, viability, lactated dehydrogenase (LDH) and nitric oxide (NO) production in endothelial progenitor cells (EPCs) under high concentration of glucose. (A, B) Flow cytometry showing the effect of SAA on the apoptosis of high-glucose (HG)-treated endothelial progenitor cells (EPCs). *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (C, D) Tube formation assay showing the effect of SAA on the tube formation of HG-treated EPCs. *** P<0.001 compared to the control group. ## P<0.01 compared to the HG group. (E) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay showing the effect of SAA on the viability of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (F) LDH assay showing the effect of SAA on the cytotoxicity of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (G) 3-Amino,4-aminomethyl-2′,7′-difluorescein diacetate (DAF-FM DA) was used to detect the effect of SAA on NO production in HG-treated EPCs. ** P<0.01 compared to the control group. # P<0.05 compared to the HG group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h) and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h).](https://jours.isi-science.com/imageXml.php?i=medscimonit-27-e928153-g002.jpg&idArt=928153&w=1000) Figure 2. Salvianic acid A (SAA) at different concentrations regulated apoptosis, tube formation, viability, lactated dehydrogenase (LDH) and nitric oxide (NO) production in endothelial progenitor cells (EPCs) under high concentration of glucose. (A, B) Flow cytometry showing the effect of SAA on the apoptosis of high-glucose (HG)-treated endothelial progenitor cells (EPCs). *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (C, D) Tube formation assay showing the effect of SAA on the tube formation of HG-treated EPCs. *** P<0.001 compared to the control group. ## P<0.01 compared to the HG group. (E) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay showing the effect of SAA on the viability of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (F) LDH assay showing the effect of SAA on the cytotoxicity of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (G) 3-Amino,4-aminomethyl-2′,7′-difluorescein diacetate (DAF-FM DA) was used to detect the effect of SAA on NO production in HG-treated EPCs. ** P<0.01 compared to the control group. # P<0.05 compared to the HG group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h) and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h).
Figure 2. Salvianic acid A (SAA) at different concentrations regulated apoptosis, tube formation, viability, lactated dehydrogenase (LDH) and nitric oxide (NO) production in endothelial progenitor cells (EPCs) under high concentration of glucose. (A, B) Flow cytometry showing the effect of SAA on the apoptosis of high-glucose (HG)-treated endothelial progenitor cells (EPCs). *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (C, D) Tube formation assay showing the effect of SAA on the tube formation of HG-treated EPCs. *** P<0.001 compared to the control group. ## P<0.01 compared to the HG group. (E) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay showing the effect of SAA on the viability of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (F) LDH assay showing the effect of SAA on the cytotoxicity of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (G) 3-Amino,4-aminomethyl-2′,7′-difluorescein diacetate (DAF-FM DA) was used to detect the effect of SAA on NO production in HG-treated EPCs. ** P<0.01 compared to the control group. # P<0.05 compared to the HG group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h) and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h). 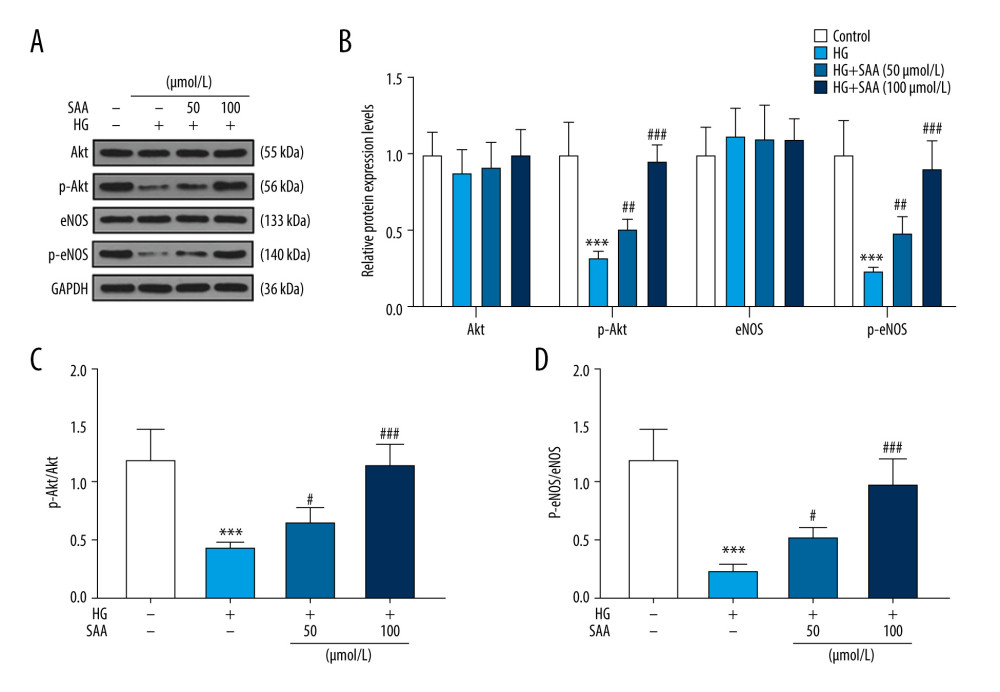 Figure 3. Effect of salvianic acid A (SAA) at different concentrations on the expression level of Akt/eNOS pathway was detected by western blotting. (A, B) Western blotting was performed to detect the protein levels of Akt, p-Akt, eNOS, and p-eNOS. *** P<0.001 compared to the control group. ## P<0.01 compared to the high-glucose (HG) group. (C) The ratio of p-Akt/Akt. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (D) The ratio of p-eNOS/eNOS. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h) and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h).
Figure 3. Effect of salvianic acid A (SAA) at different concentrations on the expression level of Akt/eNOS pathway was detected by western blotting. (A, B) Western blotting was performed to detect the protein levels of Akt, p-Akt, eNOS, and p-eNOS. *** P<0.001 compared to the control group. ## P<0.01 compared to the high-glucose (HG) group. (C) The ratio of p-Akt/Akt. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (D) The ratio of p-eNOS/eNOS. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h) and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h). 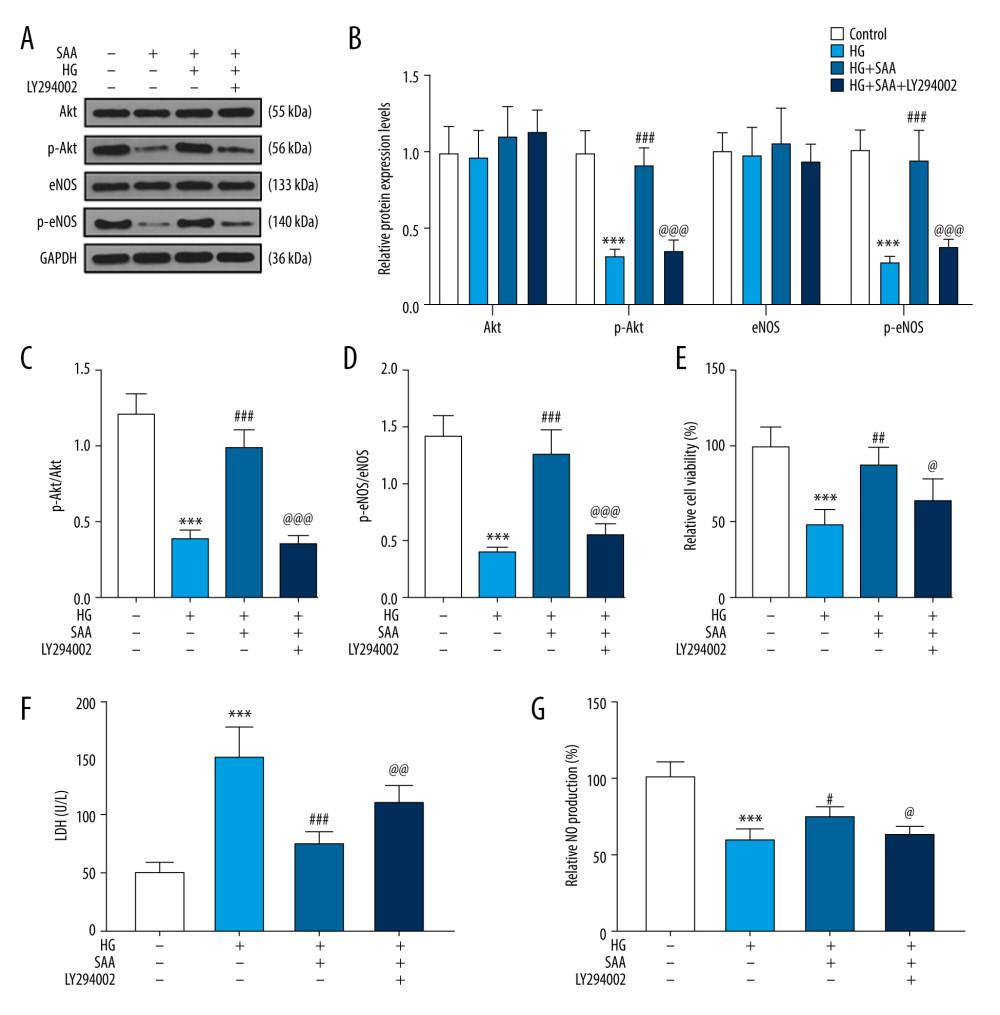 Figure 4. Effects of combined salvianic acid A (SAA) (100 μmol/L) and phosphoinositide 3-kinase (PI3K) inhibitor on the expressions of Akt/endothelial nitric oxide synthase (eNOS) pathway-related proteins, cell viability, lactated dehydrogenase (LDH) and nitric oxide (NO) production in high-glucose (HG)-treated EPCs. (A, B) the expression of Akt/eNOS pathway-related proteins was measured by western blotting. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (C) The ratio of phosphorylated (p)-Akt/Akt. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (D) The ratio of phosphorylated (p)-eNOS/eNOS. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (E) Detection of cell viability after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. ## P<0.01 compared to the HG group. @ P<0.05 compared to the HG+SAA group. (F) The levels of LDH after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@ P<0.01 compared to the HG+SAA group. (G) The level of NO production after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. @ P<0.05 compared to the HG+SAA group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+SAA group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured by 30 mmol/L glucose for 24 h) and HG+SAA+LY294002 group (cells were pretreated with 100 μmol/L SAA for 24 h and LY294002 for 1 h and then cultured with 30 mmol/L glucose for 24 h).
Figure 4. Effects of combined salvianic acid A (SAA) (100 μmol/L) and phosphoinositide 3-kinase (PI3K) inhibitor on the expressions of Akt/endothelial nitric oxide synthase (eNOS) pathway-related proteins, cell viability, lactated dehydrogenase (LDH) and nitric oxide (NO) production in high-glucose (HG)-treated EPCs. (A, B) the expression of Akt/eNOS pathway-related proteins was measured by western blotting. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (C) The ratio of phosphorylated (p)-Akt/Akt. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (D) The ratio of phosphorylated (p)-eNOS/eNOS. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (E) Detection of cell viability after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. ## P<0.01 compared to the HG group. @ P<0.05 compared to the HG+SAA group. (F) The levels of LDH after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@ P<0.01 compared to the HG+SAA group. (G) The level of NO production after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. @ P<0.05 compared to the HG+SAA group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+SAA group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured by 30 mmol/L glucose for 24 h) and HG+SAA+LY294002 group (cells were pretreated with 100 μmol/L SAA for 24 h and LY294002 for 1 h and then cultured with 30 mmol/L glucose for 24 h). 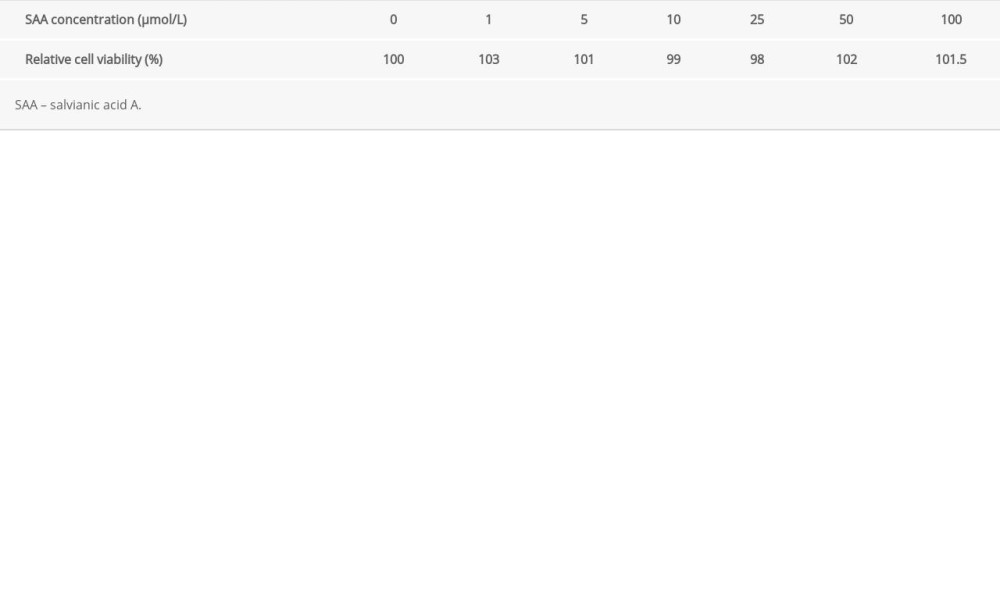
References
1. Schmidt AM, Highlighting diabetes mellitus: The epidemic continues: Arterioscler Thromb Vasc Biol, 2018; 38(1); e1-e8
2. Saeedi P, Petersohn I, Salpea P, Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition: Diabetes Res Clin Pract, 2019; 157; 107843
3. Katakami N, Mechanism of development of atherosclerosis and cardiovascular disease in diabetes mellitus: J Atheroscler Thromb, 2018; 25(1); 27-39
4. Madonna R, Pieragostino D, Balistreri CR, Diabetic macroangiopathy: Pathogenetic insights and novel therapeutic approaches with focus on high glucose-mediated vascular damage: Vascul Pharmacol, 2018 [Online ahead of print]
5. Schalkwijk CG, Stehouwer CD, Vascular complications in diabetes mellitus: The role of endothelial dysfunction: Clin Sci (Lond), 2005; 109(2); 143-59
6. Fadini GP, Sartore S, Agostini C, Avogaro A, Significance of endothelial progenitor cells in subjects with diabetes: Diabetes Care, 2007; 30(5); 1305-13
7. Chong MS, Ng WK, Chan JK, Concise review: Endothelial progenitor cells in regenerative medicine: Applications and challenges: Stem Cells Transl Med, 2016; 5(4); 530-38
8. Chopra H, Hung MK, Kwong DL, Insights into endothelial progenitor cells. Origin, classification, potentials, and prospects: Stem Cells Int, 2018; 2018 9847015
9. Phillips C, Boyd M, Assessment, management, and health implications of early-onset preeclampsia: Nurs Womens Health, 2016; 20(4); 400-14
10. Tongers J, Losordo DW, Landmesser U, Stem and progenitor cell-based therapy in ischaemic heart disease: Promise, uncertainties, and challenges: Eur Heart J, 2011; 32(10); 1197-206
11. Huang HT, Liu ZC, Wu KQ, MiR-92a regulates endothelial progenitor cells (EPCs) by targeting GDF11 via activate SMAD2/3/FAK/Akt/eNOS pathway: Ann Transl Med, 2019; 7(20); 563
12. Zhou L, Zuo Z, Chow MS, Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use: J Clin Pharmacol, 2005; 45(12); 1345-59
13. Guo J, Zhang Y, Zeng L, Salvianic acid A protects L-02 cells against gamma-irradiation-induced apoptosis via the scavenging of reactive oxygen species: Environ Toxicol Pharmacol, 2013; 35(1); 117-30
14. Zhang H, Liu YY, Jiang Q, Salvianolic acid A protects RPE cells against oxidative stress through activation of Nrf2/HO-1 signaling: Free Radic Biol Med, 2014; 69; 219-28
15. Chen YH, Lin SJ, Ku HH, Salvianolic acid B attenuates VCAM-1 and ICAM-1 expression in TNF-alpha-treated human aortic endothelial cells: J Cell Biochem, 2001; 82(3); 512-21
16. Song Q, Zhang Y, Han X, Potential mechanisms underlying the protective effects of salvianic acid A against atherosclerosis in vivo and vitro: Biomed Pharmacother, 2019; 109; 945-56
17. Krause BJ, Hanson MA, Casanello P, Role of nitric oxide in placental vascular development and function: Placenta, 2011; 32(11); 797-805
18. Rubbo H, Parthasarathy S, Barnes S, Nitric oxide inhibition of lipoxygenase-dependent liposome and low-density lipoprotein oxidation: Termination of radical chain propagation reactions and formation of nitrogen-containing oxidized lipid derivatives: Arch Biochem Biophys, 1995; 324(1); 15-25
19. Holy EW, Jakob P, Eickner T, PI3K/p110α inhibition selectively interferes with arterial thrombosis and neointima formation, but not re-endothelialization: Potential implications for drug-eluting stent design: Eur Heart J, 2014; 35(12); 808-20
20. Alegria-Schaffer A, Lodge A, Vattem K, Performing and optimizing Western blots with an emphasis on chemiluminescent detection [Chapter 33]: Guide to Protein Purification 2nd ed. Methods in Enzymology, 2009; 573-99
21. Gregg EW, Sattar N, Ali MK, The changing face of diabetes complications: Lancet Diabetes Endocrinol, 2016; 4(6); 537-47
22. Ambasta RK, Kohli H, Kumar P, Multiple therapeutic effect of endothelial progenitor cell regulated by drugs in diabetes and diabetes related disorder: J Transl Med, 2017; 15(1); 185
23. Zhao Z, Ma X, Ma J, Naringin enhances endothelial progenitor cell (EPC) proliferation and tube formation capacity through the CXCL12/CXCR4/PI3K/Akt signaling pathway: Chem Biol Interact, 2018; 286; 45-51
24. Chen YH, Lin SJ, Lin FY, High glucose impairs early and late endothelial progenitor cells by modifying nitric oxide-related but not oxidative stress-mediated mechanisms: Diabetes, 2007; 56(6); 1559-68
25. Yong L, Xei F, Xu XF, Experimental study on apoptosis of TNFR1 receptor pro-endothelial progenitor cells activated by high glucose induced oxidative stress: Int J Clin Exp Med, 2015; 8(11); 19969-81
26. Guo Y, Li Y, Xue L, Salvia miltiorrhiza: An ancient Chinese herbal medicine as a source for anti-osteoporotic drugs: J Ethnopharmacol, 2014; 155(3); 1401-16
27. Gao L, Siu PM, Chan SW, Lai CWK, Cardiovascular protective effects of salvianic acid A on db/db mice with elevated homocysteine level: Oxid Med Cell Longev, 2017; 2017 9506925
28. Zhao QT, Guo Q-M, Wang P, Wang Q, Salvianic acid A inhibits lipopolysaccharide-induced apoptosis through regulating glutathione peroxidase activity and malondialdehyde level in vascular endothelial cells: Chin J Nat Medicines, 2012; 10(1); 53-57
29. Teng F, Yin Y, Cui Y, Salvianolic acid A inhibits endothelial dysfunction and vascular remodeling in spontaneously hypertensive rats: Life Sci, 2016; 144; 86-93
30. Sun L, Zhao R, Zhang L, Salvianolic acid A inhibits PDGF-BB induced vascular smooth muscle cell migration and proliferation while does not constrain endothelial cell proliferation and nitric oxide biosynthesis: Molecules, 2012; 17(3); 3333-47
31. Saad MI, Abdelkhalek TM, Saleh MM, Insights into the molecular mechanisms of diabetes-induced endothelial dysfunction: Focus on oxidative stress and endothelial progenitor cells: Endocrine, 2015; 50(3); 537-67
32. Goligorsky MS, Vascular endothelium in diabetes: Am J Physiol Renal Physiol, 2017; 312(2); F266-75
33. Asahara T, Takahashi T, Masuda H, VEGF contributes to postnatal neovascularization by mobilizing bone marrow-derived endothelial progenitor cells: EMBO J, 1999; 18(14); 3964-72
34. Yu J, Wang Q, Wang H, Activation of liver X receptor enhances the proliferation and migration of endothelial progenitor cells and promotes vascular repair through PI3K/Akt/eNOS signaling pathway activation: Vascul Pharmacol, 2014; 62(3); 150-61
35. Cao W, Cui J, Li S, Crocetin restores diabetic endothelial progenitor cell dysfunction by enhancing NO bioavailability via regulation of PI3K/AKT-eNOS and ROS pathways: Life Sci, 2017; 181; 9-16
36. Desouza CV, Hamel FG, Bidasee K, O’Connell K, Role of inflammation and insulin resistance in endothelial progenitor cell dysfunction: Diabetes, 2011; 60(4); 1286-94
37. Murdock KW, LeRoy AS, Lacourt TE, Executive functioning and diabetes: The role of anxious arousal and inflammation: Psychoneuroendocrinology, 2016; 71; 102-9
38. Oh KS, Oh BK, Mun J, Salvianolic acid A suppress lipopolysaccharide-induced NF-kappaB signaling pathway by targeting IKKbeta: Int Immunopharmacol, 2011; 11(11); 1901-6
39. Mangialardi G, Katare R, Oikawa A, Diabetes causes bone marrow endothelial barrier dysfunction by activation of the RhoA-Rho-associated kinase signaling pathway: Arterioscler Thromb Vasc Biol, 2013; 33(3); 555-64
Figures
Figure 1. Identification of bone marrow-derived endothelial progenitor cells (EPCs) and effects of high glucose (HG) or different concentrations of salvianic acid A (SAA) on cell viability. (A) Morphology of EPCs derived from bone marrow. Scale bar=100 μm. (B) 1,1′-Dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein (Dil-ac-LDL) and fluorescein isothiocyanate-Ulex europaeus agglutinin-1 (FITC-UEA-1) double staining for identification of EPCs. Scale bar=50 μm. (C) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay was performed to determine the effects of HG (30 mmol/L) on EPC viability at 1 day, 3 days and 7 days after culture. (Forty rats were used in this experiment.) ** P<0.01 compared to the control group.Figure 2. Salvianic acid A (SAA) at different concentrations regulated apoptosis, tube formation, viability, lactated dehydrogenase (LDH) and nitric oxide (NO) production in endothelial progenitor cells (EPCs) under high concentration of glucose. (A, B) Flow cytometry showing the effect of SAA on the apoptosis of high-glucose (HG)-treated endothelial progenitor cells (EPCs). *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (C, D) Tube formation assay showing the effect of SAA on the tube formation of HG-treated EPCs. *** P<0.001 compared to the control group. ## P<0.01 compared to the HG group. (E) 3-[4,5-Dimethylthylthiazol-2-yl]-2,5 diphenyltetrazolium bromide (MTT) assay showing the effect of SAA on the viability of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (F) LDH assay showing the effect of SAA on the cytotoxicity of HG-treated EPCs. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (G) 3-Amino,4-aminomethyl-2′,7′-difluorescein diacetate (DAF-FM DA) was used to detect the effect of SAA on NO production in HG-treated EPCs. ** P<0.01 compared to the control group. # P<0.05 compared to the HG group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h) and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h).Figure 3. Effect of salvianic acid A (SAA) at different concentrations on the expression level of Akt/eNOS pathway was detected by western blotting. (A, B) Western blotting was performed to detect the protein levels of Akt, p-Akt, eNOS, and p-eNOS. *** P<0.001 compared to the control group. ## P<0.01 compared to the high-glucose (HG) group. (C) The ratio of p-Akt/Akt. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. (D) The ratio of p-eNOS/eNOS. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+50 group (cells were pretreated with 50 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h) and HG+100 group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured with 30 mmol/L glucose for 24 h).Figure 4. Effects of combined salvianic acid A (SAA) (100 μmol/L) and phosphoinositide 3-kinase (PI3K) inhibitor on the expressions of Akt/endothelial nitric oxide synthase (eNOS) pathway-related proteins, cell viability, lactated dehydrogenase (LDH) and nitric oxide (NO) production in high-glucose (HG)-treated EPCs. (A, B) the expression of Akt/eNOS pathway-related proteins was measured by western blotting. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (C) The ratio of phosphorylated (p)-Akt/Akt. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (D) The ratio of phosphorylated (p)-eNOS/eNOS. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@@ P<0.001 compared to the HG+SAA group. (E) Detection of cell viability after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. ## P<0.01 compared to the HG group. @ P<0.05 compared to the HG+SAA group. (F) The levels of LDH after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. ### P<0.001 compared to the HG group. @@ P<0.01 compared to the HG+SAA group. (G) The level of NO production after pretreatment with SAA and PI3K inhibitor. *** P<0.001 compared to the control group. # P<0.05 compared to the HG group. @ P<0.05 compared to the HG+SAA group. The cells were divided into 4 groups: control group (5.5 mmol/L glucose and 25 mmol/L mannitol), HG group (30 mmol/L glucose), HG+SAA group (cells were pretreated with 100 μmol/L SAA for 24 h and then cultured by 30 mmol/L glucose for 24 h) and HG+SAA+LY294002 group (cells were pretreated with 100 μmol/L SAA for 24 h and LY294002 for 1 h and then cultured with 30 mmol/L glucose for 24 h). In Press
07 Mar 2024 : Clinical Research
Knowledge of and Attitudes Toward Clinical Trials: A Questionnaire-Based Study of 179 Male Third- and Fourt...Med Sci Monit In Press; DOI: 10.12659/MSM.943468
08 Mar 2024 : Animal Research
Modification of Experimental Model of Necrotizing Enterocolitis (NEC) in Rat Pups by Single Exposure to Hyp...Med Sci Monit In Press; DOI: 10.12659/MSM.943443
18 Apr 2024 : Clinical Research
Comparative Analysis of Open and Closed Sphincterotomy for the Treatment of Chronic Anal Fissure: Safety an...Med Sci Monit In Press; DOI: 10.12659/MSM.944127
08 Mar 2024 : Laboratory Research
Evaluation of Retentive Strength of 50 Endodontically-Treated Single-Rooted Mandibular Second Premolars Res...Med Sci Monit In Press; DOI: 10.12659/MSM.944110
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952