09 October 2021: Review Articles
A Review of Phosphocreatine 3 Kinase δ Subtype (PI3Kδ) and Its Inhibitors in Malignancy
Qiong Xiang1ABCEF, Shuai Dong1ABDEF, Xian-Hui Li2ABCDEFG*DOI: 10.12659/MSM.932772
Med Sci Monit 2021; 27:e932772






Abstract
ABSTRACT: Most cancer deaths are caused by metastasis. The phosphocreatine 3- kinase (PI3K) family includes the I-III classes, with class I divided into 4 subtypes (α, β, γ, δ); and PI3K signaling participates in the regulatory processes of cell proliferation, differentiation, apoptosis, and glucose transport. Moreover, PI3Ks are modulators of cellular membrane lipids involved in signaling and trafficking events. The PI3Kdelta isoform (PI3Kδ), which is not only specifically expressed in hematopoietic cells, but also in different tumor cell lines, is expressed extensively. The increase in PI3Kδ activity is often associated with a variety of cancers. Currently, the strategy of tumor therapy based on PI3Kd and its related signaling pathway is developing. Besides its established role in controlling functions in autoimmunity and inflammation, the role of PI3Kδ in tumor and metastasis is not clearly elucidated, with the effects of inhibiting PI3Kδ in several types of tumors also remaining unexplored. In addition, the specific inhibitor of PI3Kδ in tumor progression and metastasis and its underlying mechanism need to be further studied. The purpose of this review is to rationalize the existing functions and mechanisms of PI3Kδ in tumor metastasis and the relationship with hematopoietic cells in cancers as well cross-talking with miRNA, which provides a new theoretical basis and potential therapeutic target for the drug therapy of tumor metastasis.
Keywords: miRNA, hematopoietic cells, PI3Kdelta, Cross-Talking, tumor metastasis, Animals, carcinogenesis, Class I Phosphatidylinositol 3-Kinases, Humans, Mice, Mutation, Neoplasms, Phosphoinositide-3 Kinase Inhibitors, Precision Medicine, Proto-Oncogene Proteins c-akt
Background
Phosphocreatine 3 kinase δ subtype (PI3Kδ), a subtype of PI3Ks, is not only mainly expressed in leukocytes [1–2], but also mediated in neurons [3] and some transformed epithelial cells [4,5]. Its high expression in the hematopoietic system is related to a variety of immune functions, mainly manifested as adaptive immunity, which plays an important role in the function of B and T cells [6], and also has an influence on mast cells [7], neutrophils [8] and macrophages [9].
PI3K is divided into 3 categories: class I, class II, and class III. Class I PI3Ks have been extensively studied and consists of a catalytic subunit p110 and a regulatory subunit (p85 or p101 heterodimer). In view of the catalytic subunit p110, class I PI3K can also be divided into PI3Kα, PI3Kβ, PI3Kδ, and PI3Kγ, whose catalytic subunits are p110α, page 110 β, page 110 δ, and p110γ, respectively. In addition, the regulatory subunit α of PI3K, PI3K company β, and PI3K company δ are p85, belonging to the RTK family, and are activated by RTKs and RAS. However, only PI3Kγ, whose regulatory subunit is P101 heterodimer, belongs to the GPCR family [10].
Most cancer deaths are caused by metastasis. Increased PI3K activity and its pathways are often associated with multiple cancers [11,12]. It is well known that the high frequency of functional mutations and amplification of the PIK3CA (p110) gene plays an important role in tumorigenesis. Alpha has been observed in human tumors. In phosphatase and tension homologous (PTEN) negative cancers, PI3Kβ is primarily involved in the production of triphosphate (3,4,5) -triphosphate, suggesting that PTEN inactivation plays a key role in tumorigenesis [13,14]. PI3Kδ and PI3Kγ participate in the immune system and inflammatory response [15,16]. However, PI3Kδ-specific inhibitors have anti-tumor effects. In addition, edilaxil (CAL101) has become the first PI3K inhibitor to be approved for the treatment of patients with recurrent chronic lymphocytic leukemia [17]. In addition, Zhao et al showed that ZSTK474, the pan-PI3K inhibitor, exhibited anti-metastatic effects by blocking the migration of PC3 cells and inhibiting the secretion of matrix metalloproteinases [18]. The role of PI3Kδ in other types of cancer such as prostate cancer metastasis has been indicated by comparing differences in DU145 cell migration and invasion after PI3K inhibitor treatment in a study by Zhang et al [19].
Increasing clinical evidence suggests that PI3K inhibitors often perform poorly in clinical trials. Due to the adverse effects of blocking other isomers, patients should be given low doses of PI3K inhibitors, which can significantly limit the clinical efficacy of PI3K inhibitors [20–22]. In this case, elucidating the role of PI3K subtype in tumor metastasis is a necessary condition for the development of specific PI3K subtype inhibitors. In the present study, the existing functions of PI3K and somatic mutations of PI3Kδ in cancer and the cross-talk with PI3K-Akt are reviewed.
Structure and Regulation of PI3Kδ
The catalytic subunit P110δPI3Kδ has an N-terminal regulatory subunit binding site, RAS-binding domain, C2 domain, and C-terminal kinase catalytic domain. The whole of class I PI3K have their regulatory subunit, P85, which includes 2 conserved Src homology-2 (SH2) domains that interplay with the phosphorylated tyrosine motif [23].
As a member of PI3K class Ia, PI3Kδ recognizes the phosphorylated tyrosine motif through the SH2 domain or is recruited into receptor complexes by targeting RAS-binding domain. The catalytic activity δ of PI3K is critical to its signal transduction and biological function [24]. While PI3Kδ is not activity, signal defects were observed in many types of cells, such as B cells, T cells, and mast cells. During signal transduction, PI3K typically responds to receptor activation that has tyrosine kinase activity. In particular, PI3Kδ also participates in the signal transduction of some GPCRs, for example, CXCR5 on B cells [25].
PI3Kδ and Cells of Hematopoietic Lineage
NK cells are an important cell type of the innate immune system. However, the mechanisms underlying the potential role of NKs in tumor immune surveillance, protection, and inhibition have not been fully elucidated. The activity of PI3Kδ appears to be related to the development of NK cells. Developmentally deficient δs and PI3Kγ expression of NK cells were observed in PI3K-deficient mice. The changes in the number of NK cells in bone marrow and peripheral blood of PI3K patients and PI3Kγ double KO mice were significantly reduced, and a wide range of immature phenotypes were observed [26,27]. They are neither deficient in extravasation of tumor growth sites nor in cytotoxicity to tumor cells [28], an observation that clearly suggests that PI3Kδ can promote metastasis by protecting tumor cells from NK cell lysis. This leads to the production of cytokines and chemokines, such as transforming growth factor-β, and downregulating the expression of NKG2D inhibits the cytotoxicity of NK cells [29].
Myeloid cells, the main type of white blood cell, are the first line of defense. Infiltration of cells into the inflammatory site is a multi-step process in which cells roll along the endothelial cells of the vascular wall under the action of selectin, attach to the endothelial cells by integrin, leave the vessel by a process of endothelial stagflation, and finally migrate through the tissue by chemical attractants. Based on its high expression in leukocytes, PI3Kδ is also a target for hematological malignancies [30], including chronic lymphocytic leukemia [31]. Indeed, PI3Kδ appears to play an important role in the late phase of cell infiltration. In vivo studies showed decreased leukocyte emigration only after the prolonged CXCL2 and TNFα treatment on the condition of PI3Kδ deficiency [32]. Moreover, leukocyte emigration was associated with the tumor cells extravasation across the endothelial barrier to distant metastatic sites [33].
Neutrophil is responsible to inflammatory mediators or pathogens, which has a critical role in pathogen clearance, however, it can also lead to injury of tissue by chronic inflammation [34]. Both of PI3Kδ and PI3Kγ are involved in this process. PIP3 accumulation is induced by fMLP in TNF-activated human neutrophils, PI3Kγ in the early stage of PIP3 production no more than 10 s of stimulation, while PI3Kδ in the late phase of PIP3 occurs within minutes of the first phase [35]. Interactions between cancer cells and neutrophils are critical to disease progression, including neutrophils infiltrating the primary tumor, neutrophils interacting with circulating tumor cells (CTC), and their involvement in the formation of pre-metastatic niches [36,37]. Identification of the axis of CTCs/neutrophils/PI3Kδ may supply novel potential targets for preventing metastasis.
Macrophages are another type of myeloid cells involved in inflammatory response, and the tumor microenvironment (TME) is an important target of tumor therapy. However, detecting and destroying tumor-promoting TMEs is a huge challenge because TMEs can have beneficial or adverse effects on tumorigenesis [38]. Similarly, M1 macrophages are well known for their anti-tumor activity; however, under the action of immunosuppressive cytokines secreted by tumor tissue or TME, they often transform into a tumor-promoting M2 phenotype [39]. In addition, many studies have shown that the microenvironment has the ability to normalize tumor cells, suggesting that re-culturing stromal cells rather than targeting tumor cells may be an effective way to treat cancer [40]. Marwick et al showed that PI3Kδ expression was upregulated in macrophages in patients with chronic obstructive pulmonary disease (COPD) and that the PI3Kδ inhibitor IC87114 restored sensitivity to glucocorticoids [41]. Environmental triggers linked to chronic inflammation, such as tobacco smoke, are not surprisingly shown to modify the local environment of tissue and contribute to angiogenesis and metastasis. It is suggested that the enhancement of PI3Kδ activity may be a potential mechanism of tumor progression, and inhibition of PI3Kδ activity may be a new way to fight tumor metastasis.
As known, mast cells are produced by precursors of bone marrow and growth factors, and PI3Kδ may play a key role in mast cells differentiation by stimulating mast cells response to varied types of growth factors [42]. In treatment of IC87114 (PI3Kδ inhibitor) in PI3Kδ KD mice or mast cells, the cytokines, including TNF and IL-6, were significantly reduced [42]. In addition, IL-6 and TNFα are usually associated with bone and marrow metastasis [43]. Similarly, IL-6 is secreted by bone marrow stromal cells and has also been shown to promote osteolysis of osteoblasts, and osteoblasts [17] have also been reported to promote metastasis of neuroblastoma [44]. These data suggest that PI3Kδ may have a critical role in genesis and progression of tumors.
PI3Kδ in Tumorigenesis and Immune Regulation
Angiogenesis is a complex, multi-step process that involves many synergistic pathways to generate stable blood vessels [45]. This process is necessary for the further growth of the primary tumor and also promotes distant metastasis and metastasis of cancer cells [46]. A series of studies have shown that targeted VEGF signaling [47,48] and the PI3K family [49] provide new prospects for angiogenesis and open up new strategies for the treatment of tumor metastasis. In particular, direct inhibition of vascular endothelial growth factor signaling [5] has proven to be very successful, but there are limitations [50,51]. PI3Ks inhibition are important for both VEGF signaling and angiogenesis, which is a potential alternative strategy [52]. Some studies have suggested that the PI3Kα subtype may play a role in tumorigenesis in cancer cells with (RAS) and PIK3CA [53–54] mutations, while the PI3Kα inhibitor BY719 and most pan-PI3K inhibitors have shown unsatisfactory results in clinical trials. Interestingly, the PI3Kδ inhibitor idelalisib, as the first specific PI3K inhibitor, has been approved by the FDA in 2014 and there is growing evidence that PI3Kδ is an attractive target for inhibition of tumor angiogenesis. PI3Kδ inhibitors negatively affected Akt and integrin b1, which are involved in cell migration and invasion [55–57] and might contribute to the anti-metastatic effect. Several PI3Kδ inhibitors, such as PI-3065 [58] and X-370 [59,60] are already in clinical trials. Above all, targeting PI3Kδ and its related pathway in endothelial cells might affect blood vessel stability and provide effective strategies for antiangiogenic therapy.
An increasing number of animal studies have shown that PI3Kδ subtype has the pharmacological effect of inhibiting tumor growth, which is not limited to hematological malignancies [61]. The explanation suggested is that these effects might be regulated by PI3Kδ in signaling pathways, which could protect tumors from immune attack [62]. Moreover, evidences from different solid tumor models indicated that the pharmacological inhibition of PI3Kδ significantly decreased metastasis [63]. These results suggested that PI3K inhibitor δ blocks blood-derived tumors and enhances the immune response to solid tumors [64].
PI3Kδ Inhibitors in Tumors
Despite significant efficacy in some solid cancers, the successful immune checkpoint blocking therapy (such as anti-PD-1) is limited by the developing mechanisms of immune resistance, such as tumor site invasion and functional CD8+ T cells development [65]. In addition, PI3Kδ inhibitors can solve these problems to inhibit immune-suppressed leukocytes, tumor-associated macrophages (TAMs) and regulatory T cells (Tregs), for example [66].
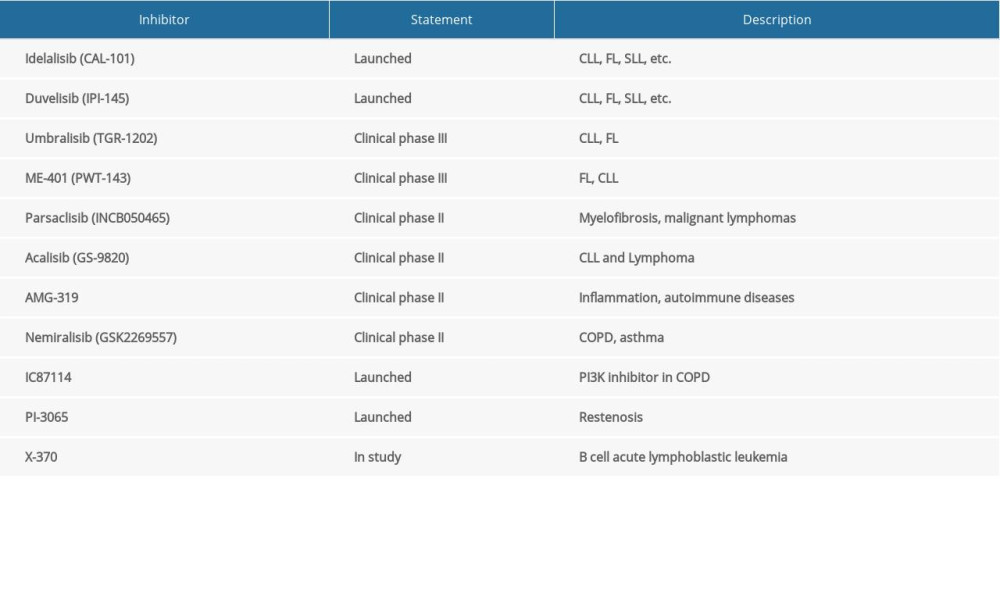
On the other hand, there are many clinical cases of acute toxicity in patients treated with PI3Kδ inhibitors (idelalisib) [67]. In addition, PI3Kδ inhibitors may lead to side effects such as inflammation, neutropenia, a high risk of infection, and death [68]. In some cases, the targeted effects of the drug can elicit a highly reactive immune response. Therefore, the administration of selective PI3K isomers at the maximum tolerated dose, in combination with other therapies, may help to overcome complications and immune system suppression [69]. Indeed, IC87114 selectively inhibits PI3Kδ with the lower IC50 concentration compared with PI3Kα, PI3Kβ, and PI3Kγ [71]. On the basis of IC87114 modification, idelalisib (CAL101) has been produced with enhanced affinity to PI3Kδ and also higher selectivity than other PI3Ks isoforms [72]. GS-9820 and AMG319 are the other 2 selective PI3Kδ inhibitors currently in phase I clinical trials, and are used to treat malignant tumors of the lymphatic system [73,74]. It has been shown that PI3Kδ inhibitors reduce tumor survival/proliferation signals by blocking PI3K component signal transduction, including Akt and ERK1/2 phosphorylation [75]. In addition, PI3Kδ inhibitors may display anti-tumor activity by inducing apoptosis in the microenvironment [76]. At the same time, evidence of PI-3065 in the treatment of solid tumors suggests that PI3K inactivation inhibits tumor growth [77]. In addition, the details mechanism of PI3Kδ inhibitors in different tumor types treatment remains to be further elucidated (Table 1).
PI3Kδ Cross-Talk with miRNAs
New evidence predicts that miRNAs regulate PI3Kδ in cancer and related signaling pathways [78,79]. MicroRNAs (miRNAs, miRs) are endogenous, non-coding RNAs, which are 18–20 nucleotides and have a role in regulating and modifying gene expression post-transcriptionally [80]. Generally, miRNAs are main and high regulators of cell behavior under normal and pathological conditions. The regulation of miRNAs can be involved in multiple stages of tumor cell diffusion from the primary site, including infiltration and exosmosis. Moreover, it is involved in tumor cell localization, tumor stromal cell interaction, dormancy and growth [81,82]. The conclusions above are supported by studies by Yuan et al on miR-26b direct targeting of PI3Kδ in human T-ALL cell lines. After treatment with PIK3CD shRNA or PIK3CD inhibitor (CAL-101), the growth of T-ALL cells was decreased and apoptosis was increased [83].
Future Challenges in PI3Kδ Research
Understanding the molecular mechanisms of PI3K δ activation of tumor progression and metastasis may provide a promising avenue for new therapeutic approaches. PI3Kδ-associated signal transduction pathways are activated in tumor cells by PI3K, and they may be cooperatively processed in the solid tumor microenvironment, resulting in reduced tumor growth. On the other hand, we need a method based on precision medicine to overcome the drug resistance mechanism of cancer cells and minimize the adverse effects and interference with blood cell homeostasis. With the development of next-generation sequencing technologies and precise drugs in clinical trials, a growing number of biomarkers predicting PI3Kδ efficacy are expected to be validated for PI3Kδ-specific tumor inhibitors.
Conclusions
In summary, the new generation of PI3Kδ provide opportunities and challenges in precision medicine and development of cancer treatments and inhibitors. As PI3Kδ advances in the cancer field each year, more and more patients will benefit from PI3Kδ-based inhibitors in the future.
References
1. Vanhaesebroeck B, Welham MJ, Kotani K, P110delta, a novel phosphoinositide 3-kinase in leukocytes: Proc Natl Acad Sci USA, 2007; 94(9); 4330-35
2. Jou ST, Chien YH, Yang YH, Identification of variations in the human phosphoinositide 3-kinase p110delta gene in children with primary B-cell immunodeficiency of unknown aetiology: Int J Immunogenet, 2006; 33(5); 361-69
3. Arimura N, Kaibuchi K, Key regulators in neuronal polarity: Neuron, 2005; 48(6); 881-84
4. De Laurentiis A, Pardo OE, Palamidessi A, The catalytic class I(A) PI3K isoforms play divergent roles in breast cancer cell migration: Cell Signal, 2011; 23(3); 529-41
5. Tzenaki N, Andreou M, Stratigi K, High levels of p110delta PI3K expression in solid tumor cells suppress PTEN activity, generating cellular sensitivity to p110delta inhibitors through PTEN activation: FASEB J, 2012; 26(6); 2498-508
6. Okkenhaug K, Bilancio A, Farjot G, Impaired B and T cell antigen receptor signaling in p110delta PI 3-kinase mutant mice: Science, 2002; 297(5583); 1031-34
7. Ali K, Bilancio A, Thomas M, Essential role for the p110delta phosphoinositide 3-kinase in the allergic response: Nature, 2004; 431(7011); 1007-11
8. Randis TM, Puri KD, Zhou H, Diacovo TG, Role of PI3Kdelta and PI3Kgamma in inflammatory arthritis and tissue localization of neutrophils: Eur J Immunol, 2008; 38(5); 1215-24
9. Horwitz SM, Koch R, Porcu P, Activity of the PI3K-δ,γ inhibitor duvelisib in a phase 1 trial and preclinical models of T-cell lymphoma: Blood, 2018; 131(8); 888-98
10. Kaneda MM, Messer KS, Ralainirina N, PI3Kγ is a molecular switch that controls immune suppression: Nature, 2016; 539(7629); 437-42
11. Fruman DA, Chiu H, Hopkins BD, The PI3K pathway in human disease: Cell, 2017; 170(4); 605-35
12. Janku F, Yap TA, Meric-Bernstam F, Targeting the PI3K pathway in cancer: Are we making headway?: Nat Rev Clin Oncol, 2018; 15(5); 273-91
13. Jia S, Liu Z, Zhang S, Essential roles of PI(3)K-p110beta in cell growth, metabolism and tumorigenesis: Nature, 2008; 454(7205); 776-79
14. Berenjeno IM, Guillermet-Guibert J, Pearce W, Both p110α and p110β isoforms of PI3K can modulate the impact of loss-of-function of the PTEN tumour suppressor: Biochem J, 2012; 442(1); 151-59
15. Fruman DA, Phosphoinositide 3-kinase and its targets in B-cell and T-cell signaling: Curr Opin Immunol, 2004; 16(3); 314-20
16. Aksoy E, Saveanu L, Manoury B, The isoform selective roles of PI3Ks in dendritic cell biology and function: Front Immunol, 2018; 15; 2574
17. Hallek M, Shanafelt TD, Eichhorst B, Chronic lymphocytic leukaemia: Lancet, 2018; 391(10129); 1524-37
18. Zhao W, Guo W, Zhou Q, In vitro antimetastatic effect of phosphatidylinositol 3-kinase inhibitor ZSTK474 on prostate cancer PC3 cells: Int J Mol Sci, 2013; 14; 13577-91
19. Zhang Z, Liu J, Wang Y, Phosphatidylinositol 3-kinase b and d isoforms play key roles in metastasis of prostate cancer DU145 cells: FASEB J, 2018; 32(11); 5967-75
20. Rodon J, Dienstmann R, Serra V, Tabernero J, Development of PI3K inhibitors: Lessons learned from early clinical trials: Nat Rev Clin Oncol, 2013; 10(3); 143-53
21. Polivka J, Janku F, Molecular targets for cancer therapy in the PI3K/AKT/mTOR pathway: Pharmacol Ther, 2014; 142(2); 164-75
22. Smith GC, Ong WK, Rewcastle GW, Effects of acutely inhibiting PI3K isoforms and mTOR on regulation of glucose metabolism in vivo: Biochem J, 2012; 442(1); 161-69
23. Marone R, Cmiljanovic V, Giese B, Wymann MP, Targeting phosphoinositide 3-kinase: Moving towards therapy: Biochim Biophys Acta, 2008; 1784(1); 159-85
24. Geering B, Cutillas PR, Nock G, Class IA phosphoinositide 3-kinases are obligate p85–p110 heterodimers: Proc Natl Acad Sci USA, 2007; 104(19); 7809-14
25. Durand CA, Hartvigsen K, Fogelstrand L, Phosphoinositide 3-kinase p110 delta regulates natural antibody production, marginal zone and B-1 B cell function, and autoantibody responses: J Immunol, 2009; 183(9); 5673-84
26. Oak JS, Fruman DA, Role of phosphoinositide 3-kinase signaling in autoimmunity: Autoimmunity, 2007; 40(6); 433-41
27. Tassi I, Cella M, Gilfillan S, p110gamma and p110delta phosphoinositide 3-kinase signaling pathways synergize to control development and functions of murine NK cells: Immunity, 2007; 27(2); 214-27
28. Saudemont A, Garçon F, Yadi H, p110gamma and p110delta isoforms of phosphoinositide 3-kinase differentially regulate natural killer cell migration in health and disease: Proc Natl Acad Sci USA, 2009; 106(14); 5795-800
29. Kopp HG, Placke T, Salih HR, Platelet-derived transforming growth factor-beta down-regulates NKG2D there by inhibiting natural killer cell antitumor reactivity: Cancer Res, 2009; 69; 7775-83
30. Greenwell IB, Flowers CR, Blum KA, Cohen JB, Clinical use of PI3K inhibitors in B-cell lymphoid malignancies: today and tomorrow: Expert Rev Anticancer Ther, 2017; 17(3); 271-79
31. Furman RR, Sharman JP, Coutre SE, Idelalisib and rituximab in relapsed chronic lymphocytic leukemia: N Engl J Med, 2014; 370(11); 997-1007
32. Liu L, Puri KD, Penninger JM, Kubes P, Leukocyte PI3Kgamma and PI3Kdelta have temporally distinct roles for leukocyte recruitment in vivo: Blood, 2007; 110(4); 1191-98
33. López-Soto A, Gonzalez S, Smyth MJ, Galluzzi L, Control of metastasis by NK cells: Cancer Cell, 2017; 32(2); 135-54
34. Sadhu C, Masinovsky B, Dick K, Essential role of phosphoinositide 3-kinase delta in neutrophil directional movement: J Immunol, 2003; 170(5); 2647-54
35. Fumagalli L, Campa CC, Germena G, Class I phosphoinositide-3-kinases and SRC kinases play a nonredundant role in regulation of adhesion-independent and -dependent neutrophil reactive oxygen species generation: J Immunol, 2013; 190(7); 3648-60
36. Saini M, Szczerba BM, Aceto N, Circulating tumor cell-neutrophil tango along the metastatic process: Cancer Res, 2019; 79(24); 6067-73
37. Swierczak A, Mouchemore KA, Hamilton JA, Anderson RL, Neutrophils: Important contributors to tumor progression and metastasis: Cancer Metastasis Rev, 2015; 34(4); 735-51
38. Quail DF, Joyce JA, Microenvironmental regulation of tumor progression and metastasis: Nat Med, 2013; 19(11); 1423-37
39. Nielsen SR, Schmid MC, Macrophages as key drivers of cancer progression and metastasis: Mediators Inflamm, 2017; 2017; 9624760
40. Smith HA, Kang Y, The metastasis-promoting roles of tumor-associated immune cells: J Mol Med (Berl), 2013; 91(4); 411-29
41. Marwick JA, Caramori G, Casolari P, A role for phosphoinositol 3-kinase delta in the impairment of glucocorticoid responsiveness in patients with chronic obstructive pulmonary disease: J Allergy Clin Immunol, 2010; 125(5); 1146-53
42. Collmann E, Bohnacker T, Marone R, Transient targeting of phosphoinositide 3-kinase acts as a roadblock in mast cells’ route to allergy: J Allergy Clin Immunol, 2013; 132(4); 959-68
43. Hjertner O, Torgersen ML, Seidel C, Hepatocyte growth factor (HGF) induces interleukin-11 secretion from osteoblasts: A possible role for HGF in myeloma-associated osteolytic bone disease: Blood, 1999; 94(11); 3883-88
44. Harmer D, Falank C, Reagan MR, Interleukin-6 interweaves the bone marrow microenvironment, bone loss, and multiple myeloma: Front Endocrinol (Lausanne), 2019; 9; 788
45. Griffioen AW, Molema G, Angiogenesis: Potentials for pharmacologic intervention in the treatment of cancer, cardiovascular diseases, and chronic inflammation: Pharmacol Rev, 2000; 52(2); 237-68
46. Lin J, Chen Y, Wei L, Ursolic acid inhibits colorectal cancer angiogenesis through suppression of multiple signaling pathways: Int J Oncol, 2013; 43(5); 1666-74
47. Zondor SD, Medina PJ, Bevacizumab: An angiogenesis inhibitor with efficacy in colorectal and other malignancies: Ann Pharmacother, 2004; 38(7–8); 1258-64
48. Brossa A, Grange C, Mancuso L, Sunitinib but not VEGF blockade inhibits cancer stem cell endothelial differentiation: Oncotarget 10, 2015; 6(13); 11295-309
49. Rauch A, Wohner N, Christophe OD, On the versatility of von Willebrand factor: Mediterr J Hematol Infect Dis, 2013; 5; e2013046
50. Dredge K, Dalgleish AG, Marriott JB, Angiogenesis inhibitors in cancer therapy: Curr Opin Investig Drugs, 2003; 4(6); 667-74
51. Bhattarai P, Hameed S, Dai Z, Recent advances in anti-angiogenic nanomedicines for cancer therapy: Nanoscale, 2018; 10(12); 5393-423
52. Mukhopadhyay D, Datta K, Multiple regulatory pathways of vascular permeability factor/vascular endothelial growth factor (VPF/VEGF) expression in tumors: Semin Cancer Biol, 2004; 14(2); 123-30
53. Zhao JJ, Liu Z, Wang L, The oncogenic properties of mutant p110alpha and p110beta phosphatidylinositol 3-kinases in human mammary epithelial cells: Proc Natl Acad Sci USA, 2005; 102(51); 18443-48
54. Ueki K, Algenstaedt P, Mauvais-Jarvis F, Kahn CR, Positive and negative regulation of phosphoinositide 3-kinase-dependent signaling pathways by three different gene products of the p85alpha regulatory subunit: Mol Cell Biol, 2000; 20(21); 8035-46
55. Ma CC, Zhang CM, Tang LQ, Liu ZP, Discovery of novel quinazolinone derivatives as high potent and selective PI3Kδ and PI3Kδ/γ inhibitors: Eur J Med Chem, 2018; 151; 9-17
56. Ciruelos Gil EM, Targeting the PI3K/AKT/mTOR pathway in estrogen receptor-positive breast cancer: Cancer Treat Rev, 2014; 40(7); 862-71
57. Shukla S, Maclennan GT, Hartman DJ, Activation of PI3K-Akt signaling pathway promotes prostate cancer cell invasion: Int J Cancer, 2007; 121(7); 1424-32
58. Bilancio A, Rinaldi B, Oliviero MA, Inhibition of p110δ PI3K prevents inflammatory response and restenosis after artery injury: Biosci Rep, 2017; 37(5); BSR20171112
59. Wang X, Zhang X, Li BS, Simultaneous targeting of PI3Kδ and a PI3Kδ-dependent MEK1/2-Erk1/2 pathway for therapy in pediatric B-cell acute lymphoblastic leukemia: Oncotarget, 2014; 5(21); 10732-44
60. Greenwell IB, Ip A, Cohen JB, PI3K Inhibitors: Understanding toxicity mechanisms and management: Oncology (Williston Park), 2017; 31(11); 821-28
61. Eramo MJ, Mitchell CA, Regulation of PtdIns(3,4,5)P3/Akt signalling by inositol polyphosphate 5-phosphatases: Biochem Soc Trans, 2016; 44(1); 240-52
62. Killock D, Immunotherapy: PI3Kδ inhibition lifts the breaks on antitumour immunity: Nat Rev Clin Oncol, 2014; 11(8); 442
63. Kingwell K, Anticancer drugs: Horizons broaden for PI3Kδ inhibitors: Nat Rev Drug Discov, 2014; 13(8); 573
64. Hirsch E, Novelli F, Cancer: Natural-born killers unleashed: Nature, 2014; 510(7505); 342-43
65. Mahoney KM, Freeman GJ, McDermott DF, The next immune-checkpoint inhibitors: PD-1/PD-L1 blockade in melanoma: Clin Ther, 2015; 37(4); 764-82
66. Uehara M, McGrath MM, Ohori S, Regulation of T cell alloimmunity by PI3Kγ and PI3Kδ: Nat Commun 16, 2017; 8(1); 951
67. Barrientos JC, Idelalisib for the treatment of chronic lymphocytic leukemia/small lymphocytic lymphoma: Future Oncol, 2016; 12(18); 2077-94
68. Weidner A, Panarelli NC, Geyer JT, Idelalisib-associated colitis: Histologic findings in 14 patients: Am J Surg Pathol, 2015; 39; 661-67
69. Curigliano G, Shah RR, Safety and tolerability of phosphatidylinositol-3-kinase (PI3K) inhibitors in oncology: Drug Saf, 2019; 42(2); 247-262
70. Shah A, Mangaonkar A, Idelalisib: A novel PI3Kδ inhibitor for chronic lymphocytic leukemia: Ann Pharmacother, 2015; 49(10); 1162-70
71. Lodola A, Bertolini S, Biagetti M, Atropisomerism and conformational equilibria: Impact on PI3Kδ inhibition of 2-((6-Amino-9H-purin-9-yl)methyl)-5-methyl-3-(o-tolyl)quinazolin-4(3H)-one (IC87114) and its conformationally restricted analogs: J Med Chem, 2017; 60(10); 4304-15
72. Yang Q, Modi P, Newcomb T, Idelalisib: First-in-class PI3K delta inhibitor for the treatment of chronic lymphocytic leukemia, small lymphocytic leukemia, and follicular lymphoma: Clin Cancer Res, 2015; 21(7); 1537-42
73. Kater AP, Tonino SH, Spiering MHOVON Lunenburg Lymphoma Phase I/II Consortium, Final results of a phase 1b study of the safety and efficacy of the PI3Kδ inhibitor acalisib (GS-9820) in relapsed/refractory lymphoid malignancies: Blood Cancer J, 2018; 8(2); 16
74. Xin M, Duan W, Feng Y, Novel 6-aryl substituted 4-pyrrolidineaminoquinazoline derivatives as potent phosphoinositide 3-kinase delta (PI3Kδ) inhibitors: Bioorg Med Chem, 2018; 26(8); 2028-40
75. Wang X, Zhang X, Li BS, Simultaneous targeting of PI3Kδ and a PI3Kδ-dependent MEK1/2-Erk1/2 pathway for therapy in pediatric B-cell acute lymphoblastic leukemia: Oncotarget 15, 2014; 5(21); 10732-44
76. Bodo J, Zhao X, Sharma A, The phosphatidylinositol 3-kinases (PI3K) inhibitor GS-1101 synergistically potentiates histone deacetylase inhibitor-induced proliferation inhibition and apoptosis through the inactivation of PI3K and extracellular signal-regulated kinase pathways: Br J Haematol, 2013; 163(1); 72-80
77. Stark AK, Sriskantharajah S, Hessel EM, Okkenhaug K, PI3K inhibitors in inflammation, autoimmunity and cancer: Curr Opin Pharmacol, 2015; 23; 82-91
78. Lu C, Shan Z, Hong J, Yang L, MicroRNA-92a promotes epithelial-mesenchymal transition through activation of PTEN/PI3K/AKT signaling pathway in non-small cell lung cancer metastasis: Int J Oncol, 2017; 51(1); 235-44
79. Palumbo T, Faucz FR, Azevedo M, Functional screen analysis reveals miR-26b and miR-128 as central regulators of pituitary somatomammotrophic tumor growth through activation of the PTEN-AKT pathway: Oncogene, 2013; 32(13); 1651-59
80. Lai EC, Micro RNAs are complementary to 3′ UTR sequence motifs that mediate negative post-transcriptional regulation: Nat Genet, 2002; 30; 363-64
81. Kucerova L, Skolekova S, Tumor microenvironment and the role of mesenchymal stromal cells: Neoplasma, 2013; 60(1); 1-10
82. Haider MT, Taipaleenmäki H, Targeting the metastatic bone microenvironment by microRNAs: Front Endocrinol (Lausanne), 2018; 27(9); 202
83. Yuan T, Yang Y, Chen J, Regulation of PI3K signaling in T-cell acute lymphoblastic leukemia: a novel PTEN/Ikaros/miR-26b mechanism reveals a critical targetable role for PIK3CD: Leukemia, 2017; 31(11); 2355-64
In Press
07 Mar 2024 : Clinical Research
Knowledge of and Attitudes Toward Clinical Trials: A Questionnaire-Based Study of 179 Male Third- and Fourt...Med Sci Monit In Press; DOI: 10.12659/MSM.943468
08 Mar 2024 : Animal Research
Modification of Experimental Model of Necrotizing Enterocolitis (NEC) in Rat Pups by Single Exposure to Hyp...Med Sci Monit In Press; DOI: 10.12659/MSM.943443
18 Apr 2024 : Clinical Research
Comparative Analysis of Open and Closed Sphincterotomy for the Treatment of Chronic Anal Fissure: Safety an...Med Sci Monit In Press; DOI: 10.12659/MSM.944127
08 Mar 2024 : Laboratory Research
Evaluation of Retentive Strength of 50 Endodontically-Treated Single-Rooted Mandibular Second Premolars Res...Med Sci Monit In Press; DOI: 10.12659/MSM.944110
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952