26 September 2021: Animal Study
Different Anesthetic Drugs Mediate Changes in Neuroplasticity During Cognitive Impairment in Sleep-Deprived Rats via Different Factors
Mengxi Shen12ABCDEF, Naqi Lian12BCE, Chengcheng Song12DE, Chao Qin12BD, Yang Yu12AE*, Yonghao Yu12ABEDOI: 10.12659/MSM.932422
Med Sci Monit 2021; 27:e932422






Abstract
BACKGROUND: Perioperative neuro-cognitive disorders (PND) are preoperative and postoperative complications of multiple nervous systems, typically manifested as decreased memory and learning ability after surgery. It was used to replace the original definition of postoperative cognitive dysfunctions (POCD) from 2018. Our previous studies have shown that sevoflurane inhalation can lead to cognitive dysfunction in Sprague-Dawley rats, but the specific mechanism is still unclear.
MATERIAL AND METHODS: Thirty-six male Sprague-Dawley rats were randomly divided into 6 groups (n=6): the SD group was given 24-h acute sleep deprivation; Sevoflurane was inhaled for 2 h in the Sevo group. Two mL propofol was injected into the tail vein of rats in the Prop group. The rats in the SD+Sevo group and SD+Prop group were deprived of sleep before intervention in the same way as before.
RESULTS: We noted significant behavioral changes in rats treated with SIK3 inhibitors or tau phosphorylation agonists before propofol injection or sevoflurane inhalation, with associated protein levels and dendritic spine density documented. Sevoflurane anesthesia-induced cognitive impairment following acute sleep deprivation was more pronounced than sleep deprivation-induced cognitive impairment alone and resulted in increased brain SIK3 levels, increased phosphorylation of total tau and tau, and decreased acetylation modifications. After using propofol, the cognitive function returned to baseline levels with a series of reversals of cognitive dysfunction.
CONCLUSIONS: These results suggest that sevoflurane inhalation via the SIK3 pathway aggravates cognitive impairment after acute sleep deprivation and that propofol anesthesia reverses the effects of sleep deprivation by affecting modifications of tau protein.
Keywords: 14-3-3 Proteins, cognitive dysfunction, Propofol, sevoflurane, SIK3 Protein, Mouse, Sleep Deprivation, Anesthetics, Inhalation, Anesthetics, Intravenous, Animals, Neuronal Plasticity
Background
Delirium occurs early after anesthesia, and another type with a long duration is called postoperative cognitive dysfunction (POCD), which is a severe postoperative neurocognitive sequela manifested as mental disorders, anxiety, personality changes, and memory disorders [1]. POCD increases the risk of postoperative complications, prolongs hospital stay, and increases mortality.
Many shreds of evidence have shown that neuroinflammation plays an essential role in the formation of POCD, which is mainly related to oxidative stress in brain tissue, blood-brain barrier injury, and neuroinflammation caused by surgical trauma [2–4]. Postoperative cognitive dysfunction occurs in about 12% of patients with good cognition before anesthesia and non-cardiac surgery [5]. Postoperative cognitive dysfunction (POCD) is generally alleviated but rarely for long periods, although plausible biological mechanisms affect protein deposition in the brain. Results of clinical studies are often unclear due to a lack of consistent definitions and test methods.
Non-memory functions (attention) are particularly associated with cognitive impairment, while psychological symptoms of anxiety and irritability are associated with physical weakness [6]. The etiology of sporadic Alzheimer disease (AD) is considered multifactorial, with environmental, biological, and genetic factors contributing to the pathogenesis of AD. General anesthetics may be such a factor and may contribute to the development and exacerbation of this neurodegenerative disease. Intraneuronal neurofibrillary tangles (NFT), composed of hyperphosphorylated and aggregated tau proteins, are among the significant neuropathological features of AD [7]. Tau pathology is essential in AD because of its strong association with cognitive dysfunction, which is part of axonal integrity, and the increase of tau indicates that neurons are damaged. The rise in nerve mercerization and tau within 48 h after surgery suggests that general anesthesia and surgery are associated with neuronal injury in the short term [8]. Studies have also found that propofol can improve the levels of Aβ-42 and tau protein in patients with liver cancer after surgery and improve the cognitive function of patients after surgery [9].
Salt-induced kinase 3 (SIK 3) is an adenosine 5′-monophosphate-activated protein kinase (AMPK) family of related kinases involved in the regulation of cell metabolism, polar remodeling, and epithelial-mesenchymal transition [10]. It is known that salt-induced kinase phosphorylates s 262 and s 356 in the Kis motif of the repeat sequence make them immunoreactive with the antibody 12 e [11], thereby phosphorylating most efficiently [12]. Activation of SIK3 aggravates mTOR expression, which activates tau protein’s abnormal phosphorylation, ultimately affecting cognitive function.
Microtubule-associated protein (tau) is abnormally hyperphosphorylated in the filamentous inclusions of cells, which can be seen in neurodegenerative diseases of dementia and significantly affect cognitive function [11]. There is increasing evidence that pathological sections can be transferred from one neuron to another via hyperphosphorylated seeds and protein uptake and co-aggregation in host neurons. Insoluble inclusion bodies formed by proteins in neurons are neuropathological features of Alzheimer disease [13]. Tau protein aggregates accumulated in the brain seem to diffuse into the whole brain through intercellular transmission processes, including pathological tau secretion and uptake, and then the template is mistakenly folded into normal tau in the receptor cells [14].
Sevoflurane, an inhalation anesthetic widely used in pediatric surgery, is considered reasonably safe and reversible. Anesthetic sevoflurane induces tau phosphorylation and cognitive impairment in juvenile rats. Its underlying mechanisms and targeted interventions remain largely unexplored [15]. Unlike propofol, sevoflurane has an affinity for a range of protein receptors in the central and peripheral nervous systems. This confounding property could serve as the basis for the functional interaction of neurons that were not related to anesthesia but have potential effects [16].
Propofol (2,6- diisopropyl phenol) is a widely used intravenous anesthetic for rapid induction and recovery from general anesthesia. As a gamma-aminobutyric acid (GABA) receptor agonist, propofol works by enhancing the inhibitory neurotransmitter GABA. Deep anesthesia induced by propofol was more protective against POCD events in clinical trials and rat models than was mild anesthesia [17].
Sleep deprivation negatively affects cognitive performance, but its exact mechanism and cognitive effect are still controversial [18]. Lack of sleep is harmful to cognitive function. The cognitive function relies on mental effort and cognitive ability compared to more automatic control processes. Evidence shows that sleep disorder and amyloid deposition work together to damage brain function of people with average cognitive ability and increase the risk of dementia in the future [19].
Night-time sleep disruption may mediate the relationship between amyloid and cognitive impairment, indicating a potential sleep-dependent mechanism that correlates amyloid burden with cognitive decline in the brain. In patients with Alzheimer disease, the source of memory deficits is, at least to some extent, the interruption of sleep patterns, which interferes with the optimal integration of previously learned declarative information [20]. However, its specific mechanism and interaction with inhalation anesthesia are unknown.
In previous studies, we found that inhalation of 2% sevoflurane for 2 h can aggravate the cognitive dysfunction induced by orthopedic surgery in aged rats, and blood-brain barrier damage may also participate in this process, but the specific mechanism is still unclear [15,21]. Propofol, a common intravenous anesthetic, exhibits neuroprotective effects after brain injury, significantly reducing the neurotoxicity accumulation and infarct size and alleviating neurological deficits [15]. In summary, the purpose of this study was to explore the interaction mechanisms of different anesthetics and sleep deprivation and their pathways leading to changes in synaptic plasticity.
Material and Methods
ANIMALS:
We purchased male SD rats (all 8-week-old and weighing 200–250 g) from Huafukang Biotechnology Co., Ltd., Beijing, China. All animals provided standard food and adequate water in a controlled environment (temperature 20–23°C, 12-h light-dark cycle). This animal experiment was authorized by the Institutional Animal Care and Use Committee of the General Hospital of Tianjin Medical University (Tianjin, China). In this experiment, the humanized endpoint was used according to the General Hospital of Tianjin Medical University’s standard operating procedures. If a rat was considered unsuitable for the next experiment, such as due to weight loss, cachexia, or difficulty eating, drinking, or walking, it was euthanized by inhalation of carbon dioxide, but no animals were sacrificed for these reasons in this study.
MEDICATION:
Sevoflurane was purchased from Hengrui Medicine Co., Ltd., put into a volatilization tank, and input into a rat anesthesia box with a 2% concentration and 60% oxygen, and measured according to the a previously described method [22]. The control group was treated with the same concentration of oxygen under the same conditions. Propofol was purchased from Xi ‘an Limongxin Pharmaceutical Co., Ltd. SIK3 inhibitors were purchased from Abcam, and tau phosphorylation agonist was purchased from Abcam.
ACUTE SLEEP DEPRIVATION MODEL:
After 1 week of acclimation to the laboratory environment, the rats were placed in a modified sleep deprivation box (a device for establishing sleep deprivation models, Patent No. ZL 2017 2 1476596.8). Based on the original level of sleep deprivation, the same environment was used in the control group. The sleep deprivation cylinder was provided with acrylic acid cylindrical sleep deprivation tables at an interval of 15 cm at the center of the cylinder along the extended shaft centerline. The lower part of the acrylic cylindrical sleep deprivation table was connected with the cylinder with a suction cup. In the experiment, rats were placed on a table, and clean water at room temperature was put into a cylinder with a water depth of 8 cm. The sleep deprivation platform was 2 cm above the water surface. At the beginning of the REM sleep stage, animals fall into the water at the instant of sleep, thus achieving the goal of sleep deprivation. In addition, the top of the cylinder was covered with a layer of wire mesh to prevent the cylinder from escaping. At the same time, a cold light source LED lamp was turned on, which completely disrupts the sleep rhythm. When rats were deprived of sleep, the curtains were closed to exclude light, and the room temperature was kept at about 22°C to eliminate temperature interference and ensure survival. During the period of acute sleep deprivation, rats had free access to drinking water and food. Deprivation time was based on a previous study [23]. Under the above conditions, acute sleep deprivation was performed for 24 h. The rats were then removed and placed in a dry cage for 1 h to begin behavioral testing. In the control group, rats were placed on acrylic cylinders with the same height and diameter of 20 cm under the same conditions, so as to ensure that the rats could rest and eliminate the environmental interference factors. After 24 h, the control group was also removed and allowed to stand for 1 h, and behavioral testing was started.
SEVOFLURANE INHALATION MODEL:
Rats in the inhalation group were placed in a ventilation box. An air inlet pipe was connected to an animal sevoflurane volatilization box (Model f 700, Nanjing Biotechnology Co., Ltd., OnStar Corp.) and an air outlet was connected to the atmospheric environment. The bottom was covered with sodium lime to absorb expired carbon dioxide and water vapor. Rats inhaled 2% sevoflurane and 60% oxygen at a flow rate of 2 L/min through the inlet pipe for 2 h. A behavioral test was performed 1 h after anesthesia. In the sleep deprivation combined with inhalation anesthesia group, the same sleep deprivation intervention was given first, and then sevoflurane was inhaled immediately to simulate preoperative sleep deprivation. Behavioral tests were performed 1 h after completion and then compared.
For inhalation gas treatment, using a previously described method [22], rats in the inhalation group (Sevo group, SD+Sevo group) were placed in a transparent square plastic box with an inlet and an outlet. The gas sensor (HY-ALERTA handheld detector model 500; Valencia, California) was set at 1.4% from start to the end of inhalation. Carbon dioxide is carried away by calcium oxide attached to the bottom.
PART ONE: DIFFERENT COGNITIVE EFFECTS CAUSED BY DIFFERENT ANESTHETICS AFTER ACUTE SLEEP DEPRIVATION:
The experimental animals were randomly divided into 6 groups, with 6 rats in each group: a control group (Con), a sevoflurane inhalation group (Sevo), a propofol intravenous anesthesia group (Prop), a sleep deprivation group (SD), a sleep deprivation combined with inhalation group (SD+Sevo), and a sleep deprivation combined with inhalation anesthesia group (SD+Prop).
The experimental model was established under the same conditions. The control group rats were placed on a large-diameter horizontal table during sleep deprivation, and 60% oxygen was inhaled during inhalation anesthesia. An exact amount of normal saline was injected through the tail vein during intravenous anesthesia. The survival rate was calculated. The rats’ survival rate in each group was observed from day 1 to day 7 after acute sleep deprivation. After the behavioral test, the other rats were subjected to PBS cardiac perfusion and bilateral hippocampal collection. The behavioral experiments included the Y maze and Morris water maze, which were used to determine rats’ cognitive level in each group.
Western blot analysis was used to detect the changes of expression levels of SIK3, tau, phosphorylated tau, acetylated tau, and PSD95 in the hippocampus of rats with cognitive dysfunction after inhalation anesthesia or intravenous anesthesia, and sleep deprivation–inhalation/intravenous anesthesia. The changes in hippocampal synaptic plasticity in the rat brain were detected by Golgi staining (Figure 1).
PART TWO. ROLE OF SIK3 IN SEVOFLURANE INHALATION AFTER ACUTE SLEEP DEPRIVATION:
The changes in cognitive behavior were detected by Y maze and Morris water maze after intraperitoneal injection of the SIK3 inhibitor YKL-05-099. The density and morphological changes of dendritic spines in rat hippocampal neurons after administration of YKL-05-099 were detected by Golgi staining. The changes in the expression of tau, phosphorylated/acetylated tau, and PSD95 in the hippocampus of rats after inhibition by the inhibitor SIK3 were determined by western blot.
PART THREE. ROLE OF TAU AND ITS MORPHOLOGY IN PROPOFOL ANESTHESIA AFTER ACUTE SLEEP DEPRIVATION:
Forskolin was used to increase tau expression and phosphorylation, followed by Y maze and Morris water maze testing to detect the cognitive level, and western blot was used to identify phosphorylated/acetylated tau expression changes and PSD95 in the rat hippocampus.
After the phosphorylation of tau protein was increased by Forskolin, the Y maze and Morris water maze were used to detect the degree of cognitive dysfunction, and western blot was used to detect the expression levels of SIK3, phosphorylated tau (Ser-404), acetylated tau (Lys-174) and PSD95 in the hippocampus of rats. The changes of hippocampal synaptic plasticity in the rat brain were detected by Golgi staining.
SURVIVAL RATES:
The survival rates of groups were estimated within 7 days, at the end of the treatment. The test was conducted twice.
Y MAZE: Y maze test was performed according to a previously described method [24]. In this maze, rodents explore new and different natural habits and it does not need animals to learn any rules to seek advantages and avoid disadvantages. It can effectively reflect the animal’s ability to recognize and memorize new and different environments. After sleep deprivation and inhalation of isoflurane for 1 h, the test started. The test included spontaneous alternating responses and spatial recognition.
The animals were placed at the end of one arm, and the sequence of animals entering each arm within 10 min was recorded. For example, alternating is defined as entering 3 arms in succession (1, 2, 3 or 1, 3, 2). The maximum alternation is the sum of −2 times of arm entry, and then calculate the percentage=actual alternation/maximum alternation×100%. Finally, the numerical values given includes actual alternation times, maximum alternation times, percentages, the total distance of animal activities, and total arm entry.
In the first experiment, 1 arm was closed and animals were allowed to explore freely in the other 2 arms for 3 min. Two hours later, the second experiment (recall stage) was carried out, in which all arms were opened, animals moved freely in 3 arms for 3 min, and the exploration time and distance in each arm were recorded, which would shorten the exploration time and distance of the memory impairment in the new arm. Finally, the parameters to be given include entering each arm and the time and distance of exploring each arm.
MORRIS WATER MAZE, MWM:
The Morris water maze test is an experiment in which experimental animals (rats and mice) are forced to swim and learn to find hidden platforms in the water, which is mainly used to test the learning and memory ability consciousness and direction consciousness (spatial positioning) of the experimental animals on spatial location. The circular rat pool had a diameter of 160 cm, a height of 50 cm, a water depth of 30 cm, a black background, and a water temperature of 23.2°C. We marked 4 equidistant points (n, e, s, and w) on the pool wall as the test’s starting point. The pool was divided into 4 quadrants, and the platform was located in the center of any quadrant (the distance between the platform and the center of the pool wall is equal). The platform is black, with a diameter of 12 cm and a height of 29 cm. Its upper surface was less than 1 cm below the water surface, which makes the platform invisible to the naked eye. Abundant reference cues (triangles, squares, circles, diamonds, and other geometric shapes placed in each quadrant) were attached around the pool and held constant to allow the rat to locate the platform. The shadow can effectively shield stray light from entering that test area. We used blinds and indirect light sources to avoid interference from light with image capture. The movement tracks of experimental animals in the Morris water maze test were extracted, and rich behavioral quantitative indicators were calculated based on the movement tracks. Behavioral statistics and analysis were performed using Anymaze software.
POSITIONING NAVIGATION TEST:
The experiment lasted 5 days, and it was planned to train 4 times a day at fixed times. At the beginning of the training, the platform was placed in the fixed quadrant, and the rats were placed into the pool in sequence from the 4 starting points of the pool wall. The video recording system recorded the time when the rats found the platform and the swimming path. After 4 training sessions, rats were placed into the water from 4 different origins (different quadrants). If the rat finds or fails to find the platform within 120 s (a delay of 120 s was recorded), the experimenter guided it to the platform and the rat was allowed to rest on the platform for 10 s before proceeding to the next experiment. The average latency of 4 training segments per day was taken as the final result of the rats.
Sometimes, before the 10- to 30-s interval expired, the rat may fall off the platform or jump into the water to continue swimming. Once this occurred, the rats were returned to the platform and retimed to allow an interval of 10–30 s. This ensured that each rat had an equal amount of time observing and obtaining spatial information after each experiment.
SPATIAL EXPLORATION TEST:
After the rats completed the last training on the 5th day, they rested for 2 h, and the platform was taken out. The rats were then put into the water from the same inlet point, and the swimming path of the rats and their time spent in the target quadrant and time needed to cross the target quadrant within 120 s were recorded. We marked the platform’s original position with a circle on the computer screen to record the time taken to traverse the original position of the platform.
WESTERN BLOTTING:
The expression levels of SIK 3, tau, phosphorylated tau, acetylated tau, and PSD 95 in the hippocampus were determined by western blot analysis.
Using preformed gel (SurePAGE™), the total protein was sampled at 8–10 μL/well, electrophoresed at 140 V for 45 min, transferred to a nitrocellulose membrane (Bio-Rad, USA), then cultured in 5% skim milk powder based on TBST solution for 1.5 h, and we used specific antibody against the following proteins: SIK 3, tau, phosphorylated tau, acetylated tau, and PSD 95.
After overnight incubation at 4°C, samples were rinsed with TBST, and then incubated at 37°C for 1 h with goat anti-mouse antibody (1: 4000, cat # 31430, Invitrogen, USA) or goat anti-rabbit antibody (1: 4000, cat # 31466, Invitrogen, USA). The nitrocellulose membrane was washed 3 times with Twin (TBST) in buffered saline, and then ECL liquid (No. 34577, Invitrogen, USA) was added dropwise, and the sample was scanned and photographed.
An image analysis system with Bio-Rad was used. The degree of expression of the goal
protein was compared with the beta-actin version by the integrated optical density target line of protein.
GOLGI STAINING:
For analysis of the hippocampus, Golgi staining was performed using the FD Fast Golgi Staining Kit (FD Neurotechnologies, Baltimore, MD) according to the manufacturer’s guidelines. Experimental animals were anesthetized by intraperitoneal injection of sevoflurane combined with 5% Avertin.
We removed the brain from the skull as soon as possible, taking care not to damage or compress the tissue. The blood on the tissue’s surface was quickly washed with Milli-Q water, and the tissue was immersed in the same volume of the infusion mixture of A and B solutions and stored at room temperature in the dark for 2 weeks. After soaking for 6 h or the next day, we replaced the saturated solution with fresh solution. We transferred the tissue to solution C, protected from light at room temperature for 72 h. We changed the solution at least once after 24 h or the following day. The tissues were cut into 200-um-thick sections with a vibrating microtome in the presence of 22% and 20% hypertonic sucrose. Samples were transferred to gelatin-coated microscope slides (Cat #PO 101) containing Solution C and recovered (provided by the kit). We pipetted any excess solution from the slide and dried it on filter paper. The sections were dried naturally in the dark at room temperature.
The sections were rinsed twice with double-distilled or Milli-Q water for 4 min each time. The sections were placed in a mixture of 1 part solution D, 1 part solution e-, and 2 parts double-distilled or Milli-Q water for 10 min. The sections were rinsed twice with distilled or Milli-Q water for 4 min each time. The sections were dehydrated for 4 min in 50%, 75%, and 95% ethanol at each concentration gradient. The sections were then dehydrated 4 times for 4 min in absolute ethanol. Samples were cleared 3 times in xylene for 4 min each time and we capped the glass with resin. The sections were then observed and photographed under an oil-immersion microscope.
ANALYSIS OF THE DENSITY AND MORPHOLOGY OF DENDRITIC SPINES:
Increasing evidence supports the involvement of neuroinflammation in the development of Alzheimer disease and further leads to changes in the plasticity of dendritic spines [25]. The density and morphology of dendritic spines from the encoded brain sections immersed in the Golgi apparatus containing the dorsal hippocampus were analyzed. The spines’ morphology and density were observed from the dendrites of the DG granular layer in the dentate gyrus and the top (radial layer) and bottom (Oriens layer) of the CA 1 pyramidal neurons.
The dendritic spines were observed and counted according to the research method of Bolam et al [22]. Neurons that met the following criteria were selected for analysis in each experimental group: (1) unclosed dendrites; (2) dark Golgi staining, consistent throughout the range of dendrites; and (3) relative separation from adjacent neuron to avoid interfering with analysis. Each neuron was analyzed with 3–5 dendritic segments, each at least 15 mm long, and each slice was analyzed with 10–11 neurons.
According to morphological characteristics, neuronal dendritic spines can be divided into the following categories: 1) Fine spines, long neck, and small head; (2) Mushrooms: prominent spines, thin neck, big head; and (3) stubby: a very short spine without a pronounced neck and stubby appearance [26,27].
To obtain the slices’ analytic image, the dendritic fragments were imaged under a Zeiss microscope with a 63× oil-immersed objective lens under bright-field illumination [28]. We calculated the linear ridge density, expressed as the number of ridges per 10 mm dendrite length, using Image J software.
STATISTICAL ANALYSIS:
All data are expressed as standard error and mean. Shapiro-Wilk normality testing was used to test whether the data were normally distributed. Unpaired
Results
SLEEP DEPRIVATION, ANESTHESIA, OR THEIR COMBINATION DID NOT SIGNIFICANTLY AFFECT SHORT-TERM SURVIVAL:
Compared with the control group, the 7-day survival rates of the SD group, SD+Sevo group, and SD+Prop group were not significantly different (P>0.05), and after inhalation anesthesia intervention, the 7-day survival rates of the Sevo group and SD+Sevo group were not significantly different (P>0.05). Only 1 of the 36 rats initially included in the experimental study died. These data indicate that sleep deprivation propofol infusion and sevoflurane inhalation intervention did not affect survival (Figure 2). We analyzed the cause of death in rats. The possible factors included the change of water temperature in the deprivation tank and the indoor temperature during deprivation, which had no significant relationship with sleep deprivation.
DIFFERENT EFFECTS OF SEVOFLURANE INHALATION AND PROPOFOL INFUSION ON COGNITIVE IMPAIRMENT AFTER ACUTE SLEEP DEPRIVATION:
We next explored the effects of various anesthetics on cognitive impairment after acute sleep deprivation. In the spontaneous alternation test, compared with the control group (n=6), the alternation rates of the SD group (n=6) and Sevo group (21.08%) were decreased by 23.86% (Figure 3A), respectively, suggesting that the number times rats entered a new arm and the time spent there were slightly decreased (Figure 3B). Notably, compared with the control group, the SD group’s alternation rate (n=6) was significantly lower (the alternation rate was reduced by 86% after the intervention) (Figure 3A), and the tendency of rats to explore new environments was reduced to various degrees (Figure 3B).
Compared to the control group, the time spent in new arm was shortened in the SD and Sevo groups, but there was no statistically significant difference between the SD and Sevo groups (Figure 3B). The time spent after entering the new arm was significantly shorter in the SD+Sevo group, indicating that the spatial memory disorder was more significant when the combined intervention was performed (Figure 3B). Surprisingly, there was no significant impairment of the Prop group’s cognitive level, but the SD+Prop group showed cognitive function comparable to that of the Control group, suggesting that propofol infusion might have lessened the cognitive impairment caused by sleep deprivation (Figure 3A, 3B).
Similar changes in cognitive levels were also clearly demonstrated in the Morris water maze test. Compared with the Con group, the change of propofol group was small but statistically significant. The acute sleep deprivation group and Sevo group had significantly increased escape latency (Figure 4B), and the alternation rate was decreased (Figure 4A). Compared with the SD group, sleep deprivation combined with sevoflurane inhalation induced more apparent cognitive impairment, and the difference was statistically significant (Figure 4A, 4B). However, the cognitive function of the SD combined with propofol group was not significantly different from that of the Con group (Figure 4A, 4B).
SEVOFLURANE PROMOTES TAU EXPRESSION AND PHOSPHORYLATION TAU BY ACTIVATING SIK3, WHICH AGGRAVATES COGNITIVE IMPAIRMENT AFTER SLEEP DEPRIVATION:
Previous studies have shown that cognitive impairment is associated with A-β fibrillary tangles, which are also the main pathological changes in Alzheimer disease. It has been proven that sleep deprivation and isoflurane inhalation can lead to cognitive impairment, mainly memory impairment [15,29]. Based on the above conclusions, we compared the expression levels of SIK 3, tau, phosphorylated tau, acetylated tau, and postsynaptic density substance PSD 95 in the bilateral hippocampus.
In the rat study, quantitative western blot analysis showed that inhalation of 2% sevoflurane for 2 h had no effect on SIK 3 expression levels, but it enhanced hippocampal tau expression in the phosphorylation/acetylation ratio of tau compared with the Control group (Figure 5A). After 24 h of acute sleep deprivation, expression of SIK 3 was increased, tau expression was increased, and the phosphorylation/acetylation ratio was increased (Figure 5A). When combined with sleep deprivation and sevoflurane anesthesia, SIK 3 levels were slightly higher than the SD group, but tau protein levels and the phosphorylation/acetylation ratio of tau protein were significantly higher compared with the single-intervention group (Figure 5A). Here, we can observe a seemingly “double” phenomenon, suggesting that if acute sleep deprivation occurs before sevoflurane inhalation anesthesia, it will aggravate the postoperative cognitive dysfunction caused by sleep deprivation, and the above mechanism may complete the process.
The change in PSD 95 may explain this problem. Compared with the Control group, the expression levels of PSD 95 in the SD group and the Sevo group were increased, and the expression levels in the SD+Sevo group were higher than those in the other 2 groups (Figure 5A).
To explore the specific mechanism by which sevoflurane inhalation aggravates cognitive impairment caused by acute sleep deprivation, we used the SIK 3 inhibitor YKL-05-099. After the rats adapted to the environment for 1 week, they were injected with normal saline as the solvent, and 1 ml of 1%YKL-05-099 dissolved in saline was injected intraperitoneally. After 1 h, each group began to receive the interventions. In the Y maze test, individuals given an intraperitoneal injection of the inhibitor showed similar levels of spontaneous alternation and spatial recognition as did Controls. The MWM test showed no damage to the cognitive level of rats in this group after inhibitor injection.
Western blot data showed that tau protein expression did not increase in the inhibitor group exposed to sevoflurane for 2 h after acute sleep deprivation, and the phosphorylation/acetylation ratio of tau protein showed the same trend (Figure 6).
These data suggest that SIK 3 inhibitors blocked elevated tau protein levels and abnormal phosphorylation of tau in the brain, ultimately protecting cognitive function.
PROPOFOL EXERTS COGNITIVE PROTECTION BY REGULATING THE EXPRESSION AND MODIFICATION OF TAU PROTEIN:
According to the results in Westing Blot test, there was no significant difference in SIK3 expression levels and previously increased tau, p-tau, was restored to high expression after application of the sleep deprivation, accompanied by down-regulation of acetylated tau and reduction of PSD-95 expression (Figure 5B).
Compared with the simple sleep deprivation group, the propofol anesthesia group after sleep deprivation showed increased alternating percentage and new arm entry times in the Y maze (Figure 3A, 3B). The Morris water maze showed that the escape latency in the sleep deprivation combined with the intravenous anesthesia group was shortened, suggesting that the long-term learning and memory ability was repaired (Figure 4).
There were no differences in protein expression levels for propofol intravenous anesthesia compared with the Control group and no significant changes in related protein levels in the SD+Prop group (Figure 5B). This contrasts with the cognitive dysfunction after acute sleep deprivation, suggesting that propofol can reverse the cognitive dysfunction caused by sleep deprivation. For PSD-95 levels, the Prop group and the SD+Prop group showed similar expression levels to the Control group with a trend consistent with the above proteins (Figure 5B).
After Forskolin injection induced hyperphosphorylation of tau in cells, western blot analysis showed that the acetylation levels of tau protein decreased and PSD-95 expression decreased, which suggested that the advantages of propofol in relieving cognitive impairment were offset by the activation of tau phosphorylation. Therefore, propofol may play a role by inhibiting the phosphorylation of tau (Figure 7).
CHANGE OF SYNAPTIC PLASTICITY AFTER ACUTE SLEEP DEPRIVATION AND DIFFERENT ANESTHESIA INTERVENTION:
We observed the changes of bilateral hippocampal neural plasticity in each group using Golgi staining. In the DG region, decreases in dendritic spines density were statistically significant in the SD and Sevo groups (Figure 8). Among the basal dendrites of CA 1 pyramidal neurons, there were no significant changes in the SD group, while the density of dendritic spines decreased after sevoflurane inhalation, and this trend was also observed in the apical dendritic spines of CA 1.
There was no significant difference in each index between the propofol group and the Control group (Figure 8). Compared with the SD group, the density of dendritic spines in DG and CA 1 regions was significantly reduced in the SD+Sevo group, suggesting that the effects of sleep deprivation and inhalation anesthesia on cognitive function ultimately changes synaptic plasticity and thus affects long-term cognitive function (Figure 8). Notably, the SD+Prop group showed a higher density of dendritic spines, and the overall trend was better than that of the sleep deprivation group, which gives us some clues that propofol anesthesia has a significant reversal effect on the cognitive impairment caused by acute sleep deprivation.
Our hypothesis was also confirmed by the results after applying SIK3 and tau phosphorylation agonist. The density of dendritic spines in the Sevo group was significantly increased after applying the SIK3 inhibitor (Figure 9). It was decreased after tail vein injection of Forskolin, which is a type of tau phosphorylated agonist, which indicates that tau agonist offset propofol’s cognitive impairment reversal effect (Figure 10).
Discussion
Postoperative cognitive dysfunction is a severe cognitive dysfunction syndrome that has been reported to be associated with neuroinflammation. It is believed that POCD can trigger similar pathways in Alzheimer disease and lead to irreversible brain pathological changes, including long-term memory impairment (cognitive impairment) [6,30]. Our previous study has shown that exposure to inhaled anesthetics during surgery increases the incidence of POCD [31]. Sleep deprivation refers to the artificial disturbance of the original sleep rhythm, shortening of insomnia time, or creation of fragmented sleep processes, which is considered to cause hyperalgesia and participate in causing cognitive impairment [32,33]. This study indicated that sleep deprivation aggravated postoperative cognitive dysfunction induced by inhaled isoflurane and confirmed the crucial role of SIK 3 in this process and its damage mechanism on hippocampal neurons in rats.
In this study, we investigated the effects of acute sleep deprivation of different durations on rat survival rate by consulting a large dataset to find the appropriate duration of acute sleep deprivation. The rats were deprived of sleep for 12 h, 24 h, 48 h, and 72 h, and the results showed that under the same environmental factors and other conditions, the mortality in the 72-h deprivation group was higher than that in the Control group by 60% and 24%, respectively. Based on the relevant studies in the same field, the ideal acute sleep deprivation duration was finally determined to be 24 h, and significant molecular biology changes were observed in subsequent experiments.
Abnormal modification of microtubule-associated protein tau is an essential factor leading to cognitive impairment [34]. To evaluate the effect of acute sleep deprivation on sevoflurane-induced severe adverse events, we examined the expression of sleep-related SIK 3 and cognitive-related tau proteins and their modified forms, including the phosphorylated form (Ser 404) and acetylated form (Lys174). In addition, some protein PSD-95, which is closely related to tau protein and may affect synaptic plasticity, was detected PSD-95 (PSD 95 rich postsynaptic scaffold protein) in glutamatergic synapses; it consists of more than 80 proteins, such as neurotransmitter receptors, ion channels, and adhesion proteins [35].
The overexpression of tau mediates the excitatory toxicity caused by E-NMDAR activation by inhibiting the phosphorylation of extracellular signal-regulated kinase, leading to the loss of hippocampal neurons [36]. PSD 95 helps to maintain synaptic stability and is involved in forming long-term memory [37] SIK 3 increases the phosphorylation level of tau protein, and the imbalance of phosphorylation/acetylation of tau protein further affects the expression of PSD 95 and finally has an irreversible effect on synaptic morphology and density [38]. In this process, acute sleep deprivation is an essential factor leading to changes in SIK 3 levels [39]. SIK 3, tau, and p-tau levels were increased, and PSD 95 levels were decreased in the SD group compared with the Control group. The changes of the above protein levels in the Sevo group were similar to those in the SD group. In the combination intervention group, sevoflurane and propofol showed different cognitive effects. Sevoflurane significantly aggravated the cognitive impairment induced by SD, and propofol provided an excellent rescue for this impairment. Therefore, propofol anesthesia can perform well in protecting the cognitive level in patients who lack sleep for various reasons before surgery.
Studies have shown that abnormal modification of tau protein is an important starting point for cognitive impairment [14]. Inhaled anesthetics such as sevoflurane accelerates tau protein’s phosphorylation, which is an essential factor of neuroprotection. Its inhibition leads to synapse reduction and plasticity changes [36,38]. In the cognitive level test, acute sleep deprivation reduced Y maze rats’ spontaneous alternation level and significantly weakened the spatial memory ability.
Besides, SIK3, tau protein modification, and synaptic-related proteins were measured to reflect the relationship between sleep, inhaled anesthesia intervention, and cognitive impairment. To evaluate whether sleep deprivation (or inhalation anesthesia) reacts to neuronal plasticity and synaptic function, Golgi staining was also performed on rats’ bilateral hippocampus in each group. These specific markers were consistent with results for sleep and cognition-related proteins. The use of SIK3 inhibitors may alleviate the above changes. The results showed that SIK3 inhibitors could reduce cognitive impairment induced by inhalation anesthesia in male SD rats by inhibiting tau protein’s abnormal phosphorylation.
We further investigated how SIK3 inhibitors protect neuronal synapses. PSD 95 nanoclusters are the basic structural units and components of excitatory synapses, and their number characterizes the size and structural diversity of synapses [40]. The density of neurons and the number of synapses were related to the expression level of PSD 95. The results showed that acute sleep deprivation combined with sevoflurane inhalation could reduce the expression of PSD 95, and this effect was more significant when these 2 interventions were combined. Significant changes in the SD+Sevo group compared with the inhalation group suggested that sleep deprivation similarly affect synaptic plasticity. Golgi staining showed that either sleep alone or inhaled anesthetics could reduce the number of neurons and observe dendritic spines’ density and morphological changes. The reduction of neurons and dendritic spines in the SD+Sevo group was significantly more substantial than that in the SD group. SIK3, a member of the salt-induced kinase family, is related to immunoinflammation and cancer [41,42]. Similarly, it has been pointed out that the salt-induced kinase family has a profound effect on tau phosphorylation [43–45]. Sleep deprivation leads to hyperphosphorylation of SIK3, affecting the further modification and regulation of tau, and reducing the expression of PSD 95 through the tau pathway, finally affecting the number of hippocampal neurons and the density of dendritic spines.
Propofol showed superior cognitive protection in this study because its main action target is tau modification, which ultimately affects synaptic plasticity. Therefore, intravenous infusion of propofol can effectively improve the long-term learning function impairment caused by acute sleep deprivation.
Among the results of our study, there is a common conclusion that the effect of sleep and inhalational anesthesia is to reduce the cognitive level. Moreover, use of intravenous propofol anesthesia may improve the cognitive impairment caused by sleep deprivation. Our previous studies found that the inhaled anesthetic sevoflurane had a significant negative impact on postoperative cognitive dysfunction through its effects on tau and its downstream signaling pathways. This study demonstrated that acute sleep deprivation caused additional impairment of cognitive function in rats after sevoflurane inhalation. It also showed that SIK 3 and its downstream signaling pathway regulate synaptic damage caused by inhalation anesthesia.
Conclusions
In conclusion, we found that a single 24-h acute sleep deprivation caused serious cognitive impairment. Sevoflurane inhalation can affect tau protein modification through SIK3 and its downstream pathways, and eventually leads to changes in synaptic plasticity, thereby aggravating the impairment of cognitive function. Propofol infusion can inhibit the abnormal hyperphosphorylation of tau protein to a certain extent to reverse the cognitive impairment caused by sleep deprivation. Therefore, we speculate that propofol anesthesia according to the patient’s situation and the application of SIK3 inhibitors may be a promising method to alleviate postoperative cognitive dysfunction caused by anesthesia.
Figures
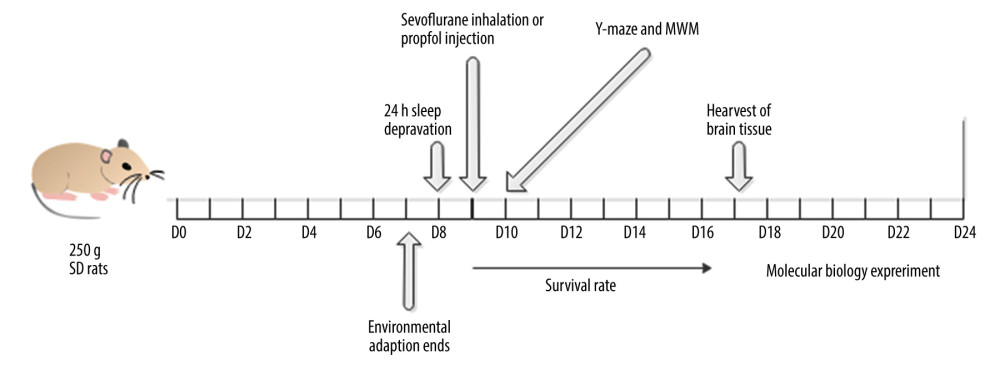 Figure 1. Experimental designSprague-Dawley male rats (6–8 weeks old, weighing 200–250 g) were randomly divided into 6 groups, with 6 rats in each group. The adaptation time was 7 days. Acute sleep deprivation lasted for 24 h from day 7, and sevoflurane inhalation lasted for 2 h from day 8. Then, the Y maze learning stage was carried out, and the test stage started on the 9th day. On the 11th day after the end of the Y maze test, the MWM test was started. After all intervention and behavior experiments, rats were killed, and the bilateral hippocampus was removed.
Figure 1. Experimental designSprague-Dawley male rats (6–8 weeks old, weighing 200–250 g) were randomly divided into 6 groups, with 6 rats in each group. The adaptation time was 7 days. Acute sleep deprivation lasted for 24 h from day 7, and sevoflurane inhalation lasted for 2 h from day 8. Then, the Y maze learning stage was carried out, and the test stage started on the 9th day. On the 11th day after the end of the Y maze test, the MWM test was started. After all intervention and behavior experiments, rats were killed, and the bilateral hippocampus was removed. 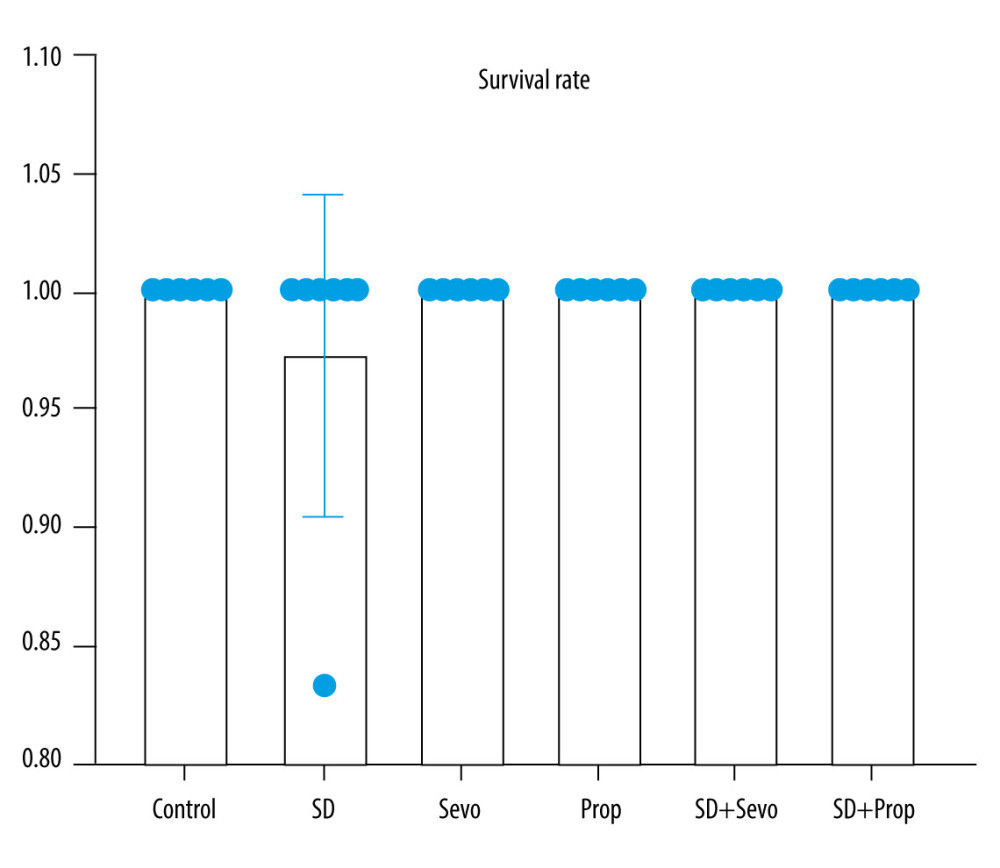 Figure 2. Rat survival rate in each groupThe 7-day survival rates of rats in the control group, the 24-h acute sleep deprivation group, the sevoflurane inhalation group, the propofol infusion group, the sleep deprivation combined with sevoflurane group, and the sleep deprivation combined with intravenous anesthesia group were observed. There was no significant difference in the survival rate among groups at different time points (P>0.05).
Figure 2. Rat survival rate in each groupThe 7-day survival rates of rats in the control group, the 24-h acute sleep deprivation group, the sevoflurane inhalation group, the propofol infusion group, the sleep deprivation combined with sevoflurane group, and the sleep deprivation combined with intravenous anesthesia group were observed. There was no significant difference in the survival rate among groups at different time points (P>0.05). 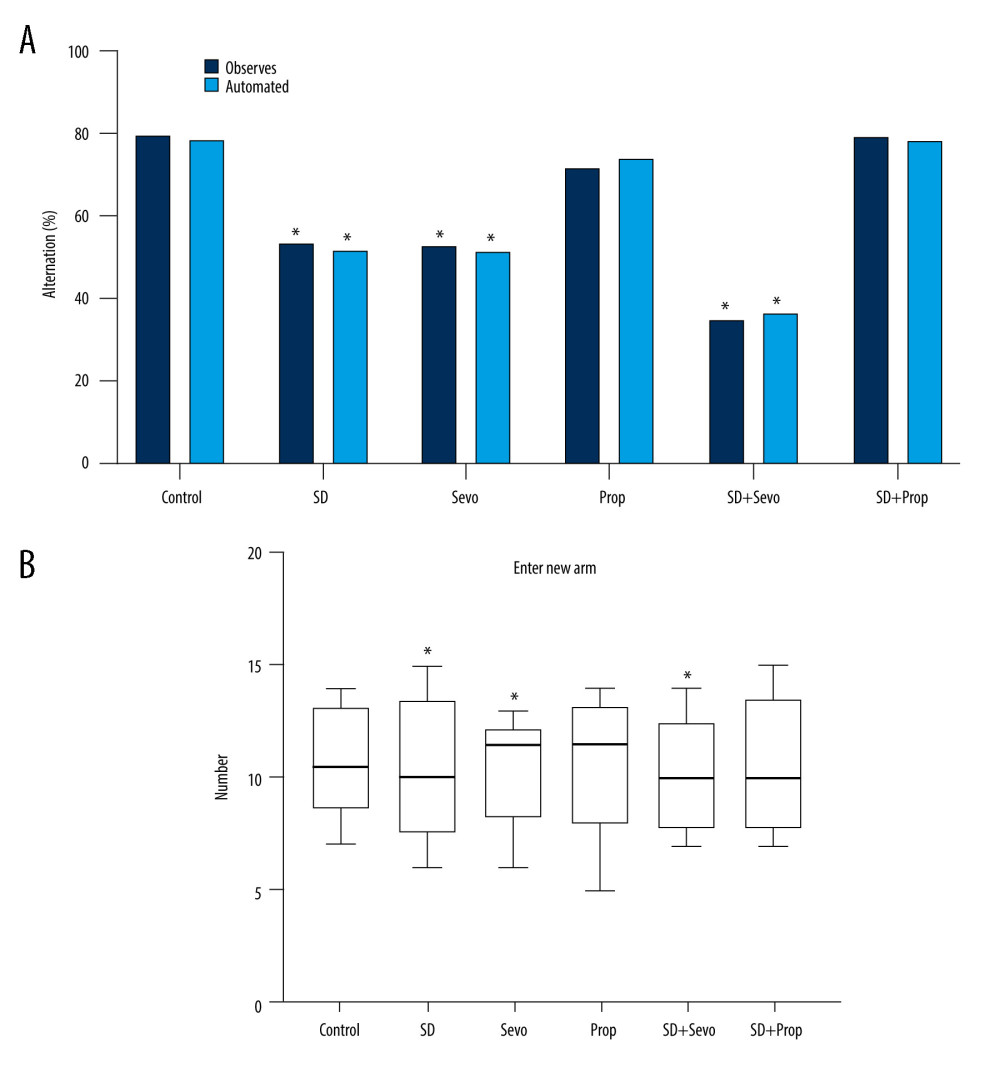 Figure 3. Behavioral changes in Y maze testing in ratsY maze test. (A) Comparison of alternating percentage between automatic Y maze and observer report. The bar graph represents the average value (n=6 for all groups). For comparison, the percentage calculated by the observer is included. Effect of treatment on alternation percentage: The results of the automatic system and the average value of observed values were analyzed independently by one-way ANOVA, and the same results were obtained (* P<0.05 Control vs SD, Control vs Sevo, SD vs SD+Sevo). (B) The number of times that rats in each group entered the new arm within 30 min (* P<0.05 Control vs SD, Control vs Sevo, SD vs SD+Sevo).
Figure 3. Behavioral changes in Y maze testing in ratsY maze test. (A) Comparison of alternating percentage between automatic Y maze and observer report. The bar graph represents the average value (n=6 for all groups). For comparison, the percentage calculated by the observer is included. Effect of treatment on alternation percentage: The results of the automatic system and the average value of observed values were analyzed independently by one-way ANOVA, and the same results were obtained (* P<0.05 Control vs SD, Control vs Sevo, SD vs SD+Sevo). (B) The number of times that rats in each group entered the new arm within 30 min (* P<0.05 Control vs SD, Control vs Sevo, SD vs SD+Sevo). 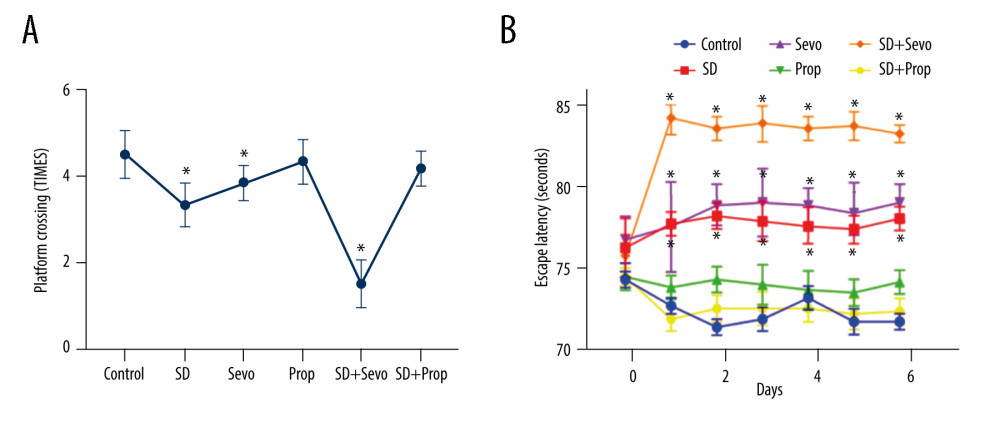 Figure 4. (A, B) Behavioral changes of MWM test in ratsMorris water maze test. Compared with the Control group, the change of propofol group was small but statistically significant. The acute sleep group and inhalation of sevoflurane group had significantly increased escape latency, and the alternation rate was decreased. Compared with the SD group, sleep deprivation combined with sevoflurane inhalation induced more apparent cognitive impairment, and the difference was statistically significant. However, the cognitive function of the SD combined with propofol group was not significantly different from that of the Control group. (* p<0.05, Control vs SD, Control vs Sevo, SD vs SD+Sevo).
Figure 4. (A, B) Behavioral changes of MWM test in ratsMorris water maze test. Compared with the Control group, the change of propofol group was small but statistically significant. The acute sleep group and inhalation of sevoflurane group had significantly increased escape latency, and the alternation rate was decreased. Compared with the SD group, sleep deprivation combined with sevoflurane inhalation induced more apparent cognitive impairment, and the difference was statistically significant. However, the cognitive function of the SD combined with propofol group was not significantly different from that of the Control group. (* p<0.05, Control vs SD, Control vs Sevo, SD vs SD+Sevo). 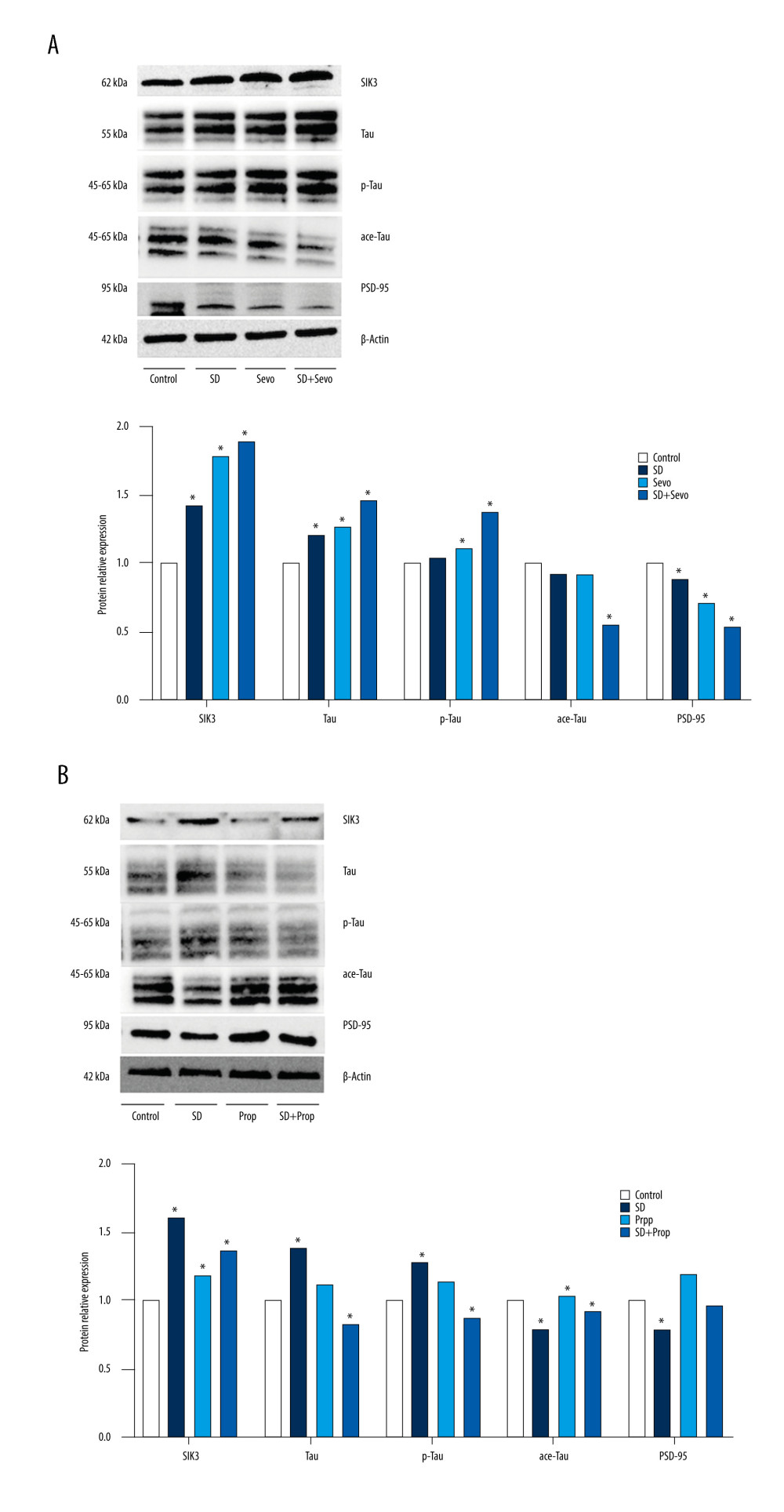 Figure 5. Expression of SIK3, PSD-95, and proteins associated with tau modification before administration of inhibitor or agonist in rats(A) Compared with the Control group, the expression level of tau protein in the hippocampus in the SD group was upregulated. Tau protein phosphorylation’s expression level was significantly upregulated, with a decreased acetylation level and an increased phosphorylation/acetylation ratio. The Sevo group exhibited increased SIK3 expression, while other changes were similar to those in the SD group. There was no statistically significant difference between the 2 groups. Compared with the SD group, the expression of SIK3 in the SD+Sevo group was upregulated. The levels of tau and its phosphorylation were increased, and the phosphorylation/acetylation ratio was increased, with statistical significance. In the change of PSD-95, the expression of the SD+Sevo group was significantly reduced. Simultaneously, the SD group and Sevo group also showed a certain degree of expression decline, and the difference was statistically significant. (* P<0.05, Control vs SD, Control vs Sevo, SD vs SD+Sevo). (B) Compared with the Con group, the expressions of SIK3, tau, p-tau were slightly increased, and ace-tau expression was slightly decreased in the SD group. The expression levels of related proteins in the Prop group were unchanged compared with the Con group. SIK3, tau, and p-tau were expressed at reduced levels. The phosphorylation/acetylation ratio was decreased in the SD+Prop group compared with the SD group. Their expression levels were similar to those in the Con group. For PSD-95 expression, SD reduced its expression, and there were no significant differences in PSD-95 expression between Con, Prop, and SD+Prop groups. (* p<0.05, Con vs SD, Con vs Prop, SD vs SD+Prop).
Figure 5. Expression of SIK3, PSD-95, and proteins associated with tau modification before administration of inhibitor or agonist in rats(A) Compared with the Control group, the expression level of tau protein in the hippocampus in the SD group was upregulated. Tau protein phosphorylation’s expression level was significantly upregulated, with a decreased acetylation level and an increased phosphorylation/acetylation ratio. The Sevo group exhibited increased SIK3 expression, while other changes were similar to those in the SD group. There was no statistically significant difference between the 2 groups. Compared with the SD group, the expression of SIK3 in the SD+Sevo group was upregulated. The levels of tau and its phosphorylation were increased, and the phosphorylation/acetylation ratio was increased, with statistical significance. In the change of PSD-95, the expression of the SD+Sevo group was significantly reduced. Simultaneously, the SD group and Sevo group also showed a certain degree of expression decline, and the difference was statistically significant. (* P<0.05, Control vs SD, Control vs Sevo, SD vs SD+Sevo). (B) Compared with the Con group, the expressions of SIK3, tau, p-tau were slightly increased, and ace-tau expression was slightly decreased in the SD group. The expression levels of related proteins in the Prop group were unchanged compared with the Con group. SIK3, tau, and p-tau were expressed at reduced levels. The phosphorylation/acetylation ratio was decreased in the SD+Prop group compared with the SD group. Their expression levels were similar to those in the Con group. For PSD-95 expression, SD reduced its expression, and there were no significant differences in PSD-95 expression between Con, Prop, and SD+Prop groups. (* p<0.05, Con vs SD, Con vs Prop, SD vs SD+Prop). 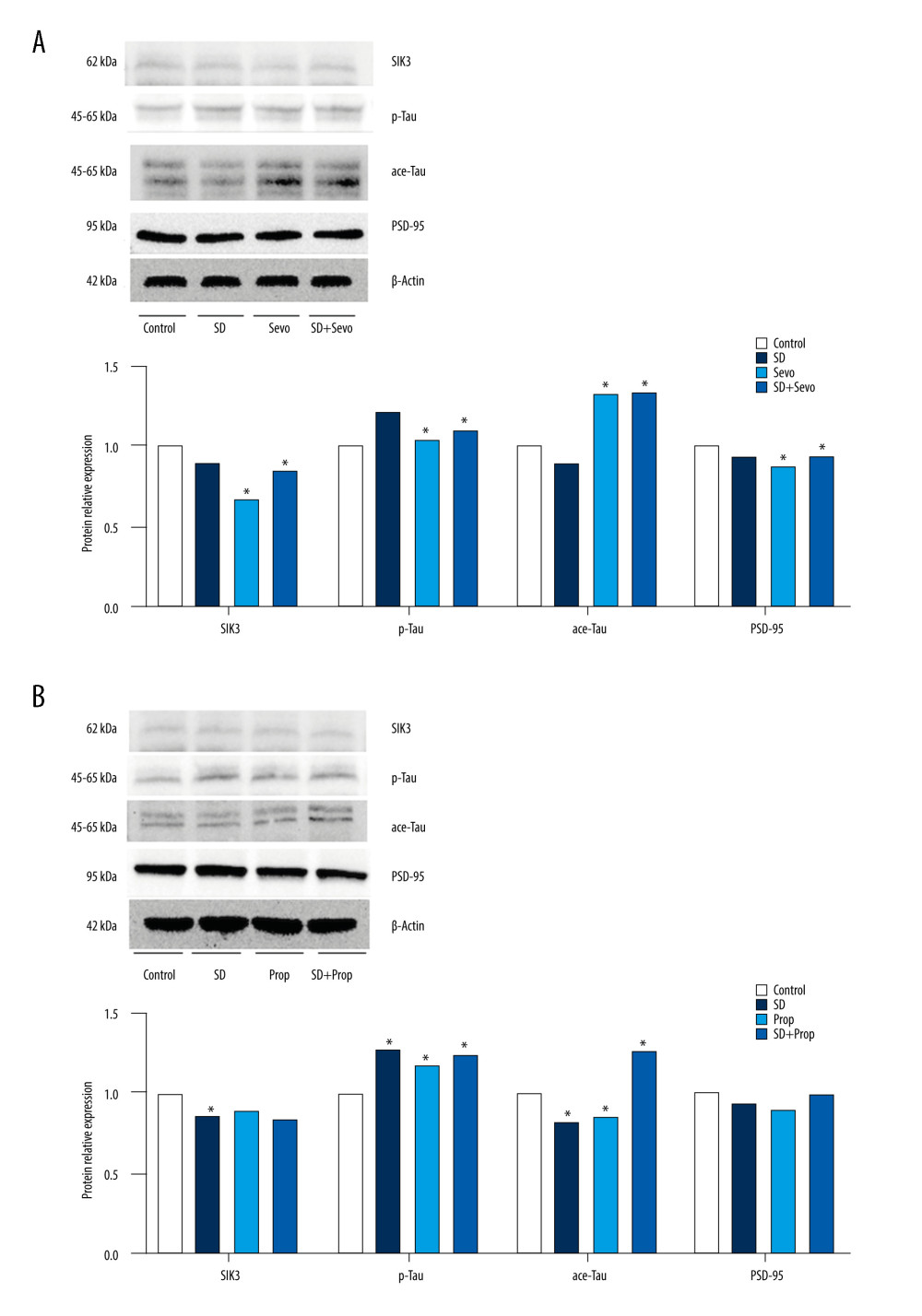 Figure 6. (A, B) Expression of SIK3, PSD-95 and proteins associated with tau modification after administration of SIK3 inhibitor in ratsIn the Control group, the expression level of tau protein in the hippocampus of the SD group was upregulated (P<0.05), the expression level of tau protein phosphorylation (Ser404) was significantly upregulated (P<0.05), and the acetylation level was decreased (P<0.05), together with the decrease in postsynaptic PSD95. There was no significant difference in each protein’s levels between the Sevo group and the SD group (P>0.05). The SD+Sevo group showed a stronger trend than the SD group (P<0.05) (n=6/group). (* P<0.05, Con vs SD, Con vs Sevo, SD vs SD+Sevo).
Figure 6. (A, B) Expression of SIK3, PSD-95 and proteins associated with tau modification after administration of SIK3 inhibitor in ratsIn the Control group, the expression level of tau protein in the hippocampus of the SD group was upregulated (P<0.05), the expression level of tau protein phosphorylation (Ser404) was significantly upregulated (P<0.05), and the acetylation level was decreased (P<0.05), together with the decrease in postsynaptic PSD95. There was no significant difference in each protein’s levels between the Sevo group and the SD group (P>0.05). The SD+Sevo group showed a stronger trend than the SD group (P<0.05) (n=6/group). (* P<0.05, Con vs SD, Con vs Sevo, SD vs SD+Sevo). 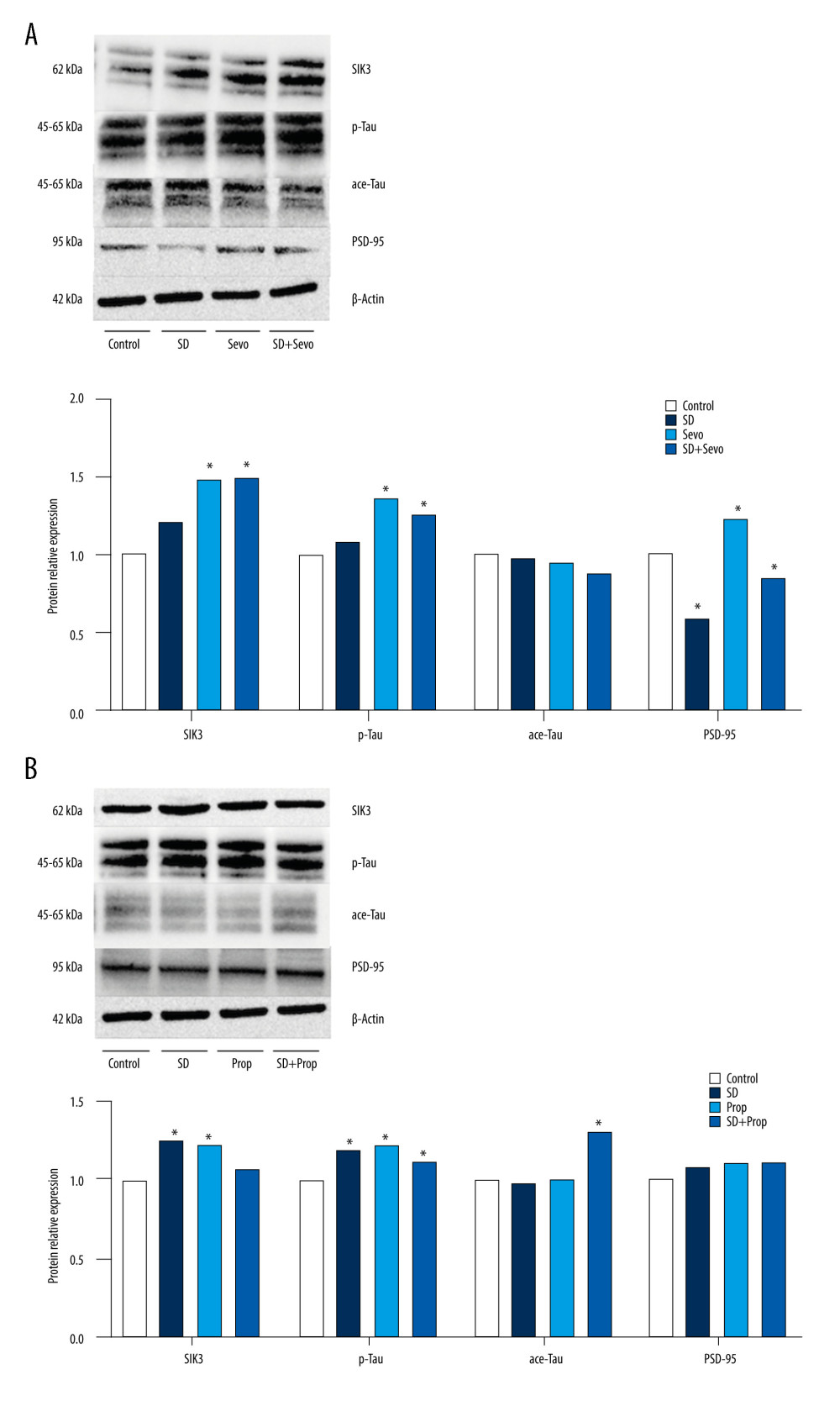 Figure 7. (A, B) Expression of SIK3, PSD-95 and proteins associated with tau modification after administration of tau agonist in ratsIntravenous anesthesia with propofol reversed cognitive impairment caused by acute sleep deprivation. In the Control group, the expression level of tau protein in the hippocampus of the SD group was upregulated (P<0.05), the expression level of tau protein phosphorylation (Ser404) was significantly upregulated (P<0.05), and the acetylation level was decreased (P<0.05), together with the decrease in postsynaptic PSD95. There was no significant difference in each protein’s levels between the Prop group and the Control group (P>0.05). The SD+Prop group outperformed the SD group in cognition, and the results were statistically significant (* P<0.05 Con vs SD, Control vs Prop, and SD vs SD+Prop).
Figure 7. (A, B) Expression of SIK3, PSD-95 and proteins associated with tau modification after administration of tau agonist in ratsIntravenous anesthesia with propofol reversed cognitive impairment caused by acute sleep deprivation. In the Control group, the expression level of tau protein in the hippocampus of the SD group was upregulated (P<0.05), the expression level of tau protein phosphorylation (Ser404) was significantly upregulated (P<0.05), and the acetylation level was decreased (P<0.05), together with the decrease in postsynaptic PSD95. There was no significant difference in each protein’s levels between the Prop group and the Control group (P>0.05). The SD+Prop group outperformed the SD group in cognition, and the results were statistically significant (* P<0.05 Con vs SD, Control vs Prop, and SD vs SD+Prop).  Figure 8. (A, B) Change in hippocampal dendritic spines before administration of inhibitors or agonistsIn the DG region, the density of dendritic spines decreased significantly in the SD and Sevo groups. There was no noticeable change in the SD group in the basal dendrites of CA 1 pyramidal neurons. However, the density of dendritic spines decreased after inhalation of sevoflurane. This trend also existed in the apical dendritic spines of CA 1. There was no significant difference between the propofol group and the Control group. The SD group density of dendritic spines in the DG and CA 1 area in the SD+Sevo group decreased significantly, suggesting that sleep deprivation and inhalation anesthesia on cognitive function eventually changed synaptic plasticity, thus affecting long-term cognitive function. It is worth noting that the SD+Prop group showed a higher density of dendritic spines. The overall trend was better than that of the sleep deprivation group, which suggests that propofol anesthesia can significantly reverse the cognitive impairment caused by acute sleep deprivation.
Figure 8. (A, B) Change in hippocampal dendritic spines before administration of inhibitors or agonistsIn the DG region, the density of dendritic spines decreased significantly in the SD and Sevo groups. There was no noticeable change in the SD group in the basal dendrites of CA 1 pyramidal neurons. However, the density of dendritic spines decreased after inhalation of sevoflurane. This trend also existed in the apical dendritic spines of CA 1. There was no significant difference between the propofol group and the Control group. The SD group density of dendritic spines in the DG and CA 1 area in the SD+Sevo group decreased significantly, suggesting that sleep deprivation and inhalation anesthesia on cognitive function eventually changed synaptic plasticity, thus affecting long-term cognitive function. It is worth noting that the SD+Prop group showed a higher density of dendritic spines. The overall trend was better than that of the sleep deprivation group, which suggests that propofol anesthesia can significantly reverse the cognitive impairment caused by acute sleep deprivation.  Figure 9. Change in hippocampal dendritic spines after administration of the SIK3 inhibitor YKL-05-099The changes of bilateral hippocampal neural plasticity in each group were observed by Golgi staining. In the DG region, there was a statistically significant increase in dendritic spine density in the SD and Sevo groups compared to before inhibitor administration. The density of dendritic spines in DG and CA 1 regions in the SD+Sevo group was slightly decreased, suggesting that the effects of sleep deprivation and inhalation anesthesia on cognitive function ultimately changed synaptic plasticity and thus affected long-term cognitive function.
Figure 9. Change in hippocampal dendritic spines after administration of the SIK3 inhibitor YKL-05-099The changes of bilateral hippocampal neural plasticity in each group were observed by Golgi staining. In the DG region, there was a statistically significant increase in dendritic spine density in the SD and Sevo groups compared to before inhibitor administration. The density of dendritic spines in DG and CA 1 regions in the SD+Sevo group was slightly decreased, suggesting that the effects of sleep deprivation and inhalation anesthesia on cognitive function ultimately changed synaptic plasticity and thus affected long-term cognitive function.  Figure 10. Change in hippocampal dendritic spines after administration of Tau modification agonistIn the DG region, dendritic spines’ density was decreased in the SD and Prop groups compared with before. The dendritic spines in DG and CA 1 regions in the SD+Prop group were denser than those in the Control group, suggesting that the effects of sleep deprivation and intravenous anesthesia on cognitive function can eventually change synaptic plasticity, thus affecting long-term cognitive function. Propofol infusion had a definite reversal effect on cognitive dysfunction caused by sleep deprivation.
Figure 10. Change in hippocampal dendritic spines after administration of Tau modification agonistIn the DG region, dendritic spines’ density was decreased in the SD and Prop groups compared with before. The dendritic spines in DG and CA 1 regions in the SD+Prop group were denser than those in the Control group, suggesting that the effects of sleep deprivation and intravenous anesthesia on cognitive function can eventually change synaptic plasticity, thus affecting long-term cognitive function. Propofol infusion had a definite reversal effect on cognitive dysfunction caused by sleep deprivation. References
1. Yoshida H, Goedert M, Phosphorylation of microtubule-associated protein tau by AMPK-related kinases: J Neurochem, 2012; 120(1); 165-76
2. Ren JG, Seth P, Clish CB, Knockdown of malic enzyme 2 suppresses lung tumor growth, induces differentiation and impacts PI3K/AKT signaling: Sci Rep, 2014; 4; 5414
3. Broadhead MJ, Horrocks MH, Zhu F, PSD95 nanoclusters are postsynaptic building blocks in hippocampus circuits: Sci Rep, 2016; 6; 24626
4. Frank RAW, Zhu F, Komiyama NH, Grant SGN, Hierarchical organization and genetically separable subfamilies of PSD95 postsynaptic supercomplexes: J Neurochem, 2017; 142(4); 504-11
5. Lombardi MS, Gilliéron C, Berkelaar M, Gabay C, Salt-inducible kinases (SIK) inhibition reduces RANKL-induced osteoclastogenesis: PLoS One, 2017; 12(10); e0185426
6. Majnarić LT, Bekić S, Babič F, Cluster analysis of the associations among physical frailty, cognitive impairment and mental disorders: Med Sci Monit, 2020; 26; e924281
7. Whittington RA, Bretteville A, Dickler MF, Planel E, Anesthesia and tau pathology: Prog Neuropsychopharmacol Biol Psychiatry, 2013; 47; 147-55
8. Evered L, Silbert B, Scott DA, Association of changes in plasma neurofilament light and Tau levels with anesthesia and surgery: Results from the CAPACITY and ARCADIAN Studies: JAMA Neurol, 2018; 75(5); 542-47
9. Hou JF, Xiao CL, Effect of propofol and sevoflurane anesthesia on postoperative cognitive function and levels of Aβ-42 and Tau in patients undergoing hepatectomy: Eur Rev Med Pharmacol Sci, 2019; 23(2); 849-56
10. Honda T, Fujiyama T, Miyoshi C, A single phosphorylation site of SIK3 regulates daily sleep amounts and sleep need in mice: Proc Natl Acad Sci USA, 2018; 115(41); 10458-63
11. Subramanian J, Michel K, Benoit M, Nedivi E, CPG15/Neuritin mimics experience in selecting excitatory synapses for stabilization by facilitating PSD95 recruitment: Cell Rep, 2019; 28(6); 1584-95.e5
12. Goedert M, Klug A, Crowther RA, Tau protein, the paired helical filament and Alzheimer’s disease: J Alzheimers Dis, 2006; 9(3 Suppl); 195-207
13. Sun XY, Tuo QZ, Liuyang ZY, Extrasynaptic NMDA receptor-induced tau overexpression mediates neuronal death through suppressing survival signaling ERK phosphorylation: Cell Death Dis, 2016; 7(11); e2449
14. Ossenkoppele R, Lyoo CH, Jester-Broms J, Assessment of demographic, genetic, and imaging variables associated with brain resilience and cognitive resilience to pathological Tau in patients with Alzheimer sisease: JAMA Neurol, 2020; 77(5); 632-42
15. Boros BD, Greathouse KM, Gentry EG, Dendritic spines provide cognitive resilience against Alzheimer’s disease: Ann Neurol, 2017; 82(4); 602-14
16. Sun H, Zhang G, Ai B, A systematic review: comparative analysis of the effects of propofol and sevoflurane on postoperative cognitive function in elderly patients with lung cancer: BMC Cancer, 2019; 19(1); 1248
17. Varnäs K, Finnema SJ, Johnström P: Br J Anaesth, 2021; 126(1); 238-44
18. Schulz K, Korz V, Hippocampal testosterone relates to reference memory performance and synaptic plasticity in male rats: Front Behav Neurosci, 2010; 4; 187
19. Staffe AT, Bech MW, Clemmensen SLK, Total sleep deprivation increases pain sensitivity, impairs conditioned pain modulation and facilitates temporal summation of pain in healthy participants: PLoS One, 2019; 14(12); e0225849
20. Benvenuti SM, Psychobiological mechanisms underlying cognitive decline in cardiac surgery patients: PhD Thesis, 2012, Padova University
21. Chakravarthy S, Saiepour MH, Bence M, Postsynaptic TrkB signaling has distinct roles in spine maintenance in adult visual cortex and hippocampus: Proc Natl Acad Sci USA, 2006; 103(4); 1071-76
22. Bolam JP, Clarke DJ, Smith AD, Somogyi P, A type of aspiny neuron in the rat neostriatum accumulates [3H]gamma-aminobutyric acid: Combination of Golgi-staining, autoradiography, and electron microscopy: J Comp Neurol, 1983; 213(2); 121-34
23. Talan J, At the Bench-Alzheimer’s disease: Unique signature in dendritic spines may explain cognitive resilience to Alzheimer’s disease: Neurology Today, 2017; 17(21); 1
24. Lv G, Li C, Wang W, Silencing SP1 alleviated sevoflurane-induced POCD development via cholinergic anti-inflammatory pathway: Neurochem Res, 2020; 45(9); 2082-90
25. Pepino MY, Eisenstein SA, Bischoff AN, Sweet dopamine: Sucrose preferences relate differentially to striatal D2 receptor binding and age in obesity: Diabetes, 2016; 65(9); 2618-23
26. Zhao G, Yang L, Wang S, TREK-2 mediates the neuroprotective effect of isoflurane preconditioning against acute cerebral ischemia in the rat: Rejuvenation Res, 2019; 22(4); 325-34
27. Yu L, Sun H, Yao L, Feng Y, An approach to using central pupils as a clinical sign to assess depth of anesthesia in infants undergoing fundus examination with inhalation of sevoflurane: J Clin Anesth, 2016; 29; 5-9
28. Wendler E, de Souza CP, Dornellas APS, Mania-like elevated mood in rats: Enhanced 50-kHz ultrasonic vocalizations after sleep deprivation: Prog Neuropsychopharmacol Biol Psychiatry, 2019; 88; 142-50
29. Fan Y, Du L, Fu Q, Inhibiting the NLRP3 inflammasome with MCC950 ameliorates isoflurane-induced pyroptosis and cognitive impairment in aged mice: Front Cell Neurosci, 2018; 12; 426
30. Koyanagi Y, Oi Y, Kobayashi M, Fast-spiking interneurons contribute to propofol-induced facilitation of firing synchrony in pyramidal neurons of the rat insular cortex: Anesthesiology, 2021; 134(2); 219-33
31. Hu N, Guo D, Wang H, Involvement of the blood-brain barrier opening in cognitive decline in aged rats following orthopedic surgery and high concentration of sevoflurane inhalation: Brain Res, 2014; 1551; 13-24
32. Henry M, Ross IL, Wolf PSA, Thomas KGF, Impaired quality and efficiency of sleep impairs cognitive functioning in Addison’s disease: Psychoneuroendocrinology, 2017; 78; 237-45
33. You JC, Jones E, Cross DE, Association of β-amyloid burden with sleep dysfunction and cognitive impairment in elderly individuals with cognitive disorders: JAMA Netw Open, 2019; 2(10); e1913383
34. Kusztor A, Raud L, Juel BE, Sleep deprivation differentially affects subcomponents of cognitive control: Sleep, 2019; 42(4); zsz016
35. Shi J, Zou X, Jiang K, Wang F, SIRT1 mediates improvement of cardiac surgery-induced postoperative cognitive dysfunction via the TLR4/NF-κB pathway: World J Biol Psychiatry, 2020; 21(10); 757-65
36. Merezhko M, Brunello CA, Yan X, Secretion of Tau via an unconventional non-vesicular mechanism: Cell Rep, 2018; 25(8); 2027-35.e4
37. Schlegel K, Awwad K, Heym RG, N368-Tau fragments generated by legumain are detected only in trace amount in the insoluble Tau aggregates isolated from AD brain: Acta Neuropathol Commun, 2019; 7(1); 177
38. Dai CL, Hu W, Tung YC, Tau passive immunization blocks seeding and spread of Alzheimer hyperphosphorylated Tau-induced pathology in 3×Tg-AD mice: Alzheimers Res Ther, 2018; 10(1); 13
39. Thornton C, Bright NJ, Sastre M, AMP-activated protein kinase (AMPK) is a tau kinase, activated in response to amyloid β-peptide exposure: Biochem J, 2011; 434(3); 503-12
40. Ponnusamy L, Kothandan G, Manoharan R, Berberine and Emodin abrogates breast cancer growth and facilitates apoptosis through inactivation of SIK3-induced mTOR and Akt signaling pathway: Biochim Biophys Acta Mol Basis Dis, 2020; 1866(11); 165897
41. Han C, Fu R, Lei W, Beneficial effects of dexmedetomidine on early postoperative cognitive dysfunction in pediatric patients with tonsillectomy: Exp Ther Med, 2018; 16(1); 420-26
42. Xiong B, Shi Q, Fang H, Dexmedetomidine alleviates postoperative cognitive dysfunction by inhibiting neuron excitation in aged rats: Am J Transl Res, 2016; 8(1); 70-80
43. O’Gara BP, Mueller A, Gasangwa DVI, Prevention of early postoperative decline: A randomized, controlled feasibility trial of perioperative cognitive training: Anesth Analg, 2020; 130(3); 586-95
44. Chen Y, Zhang Y, Ye G, Knockdown of lncRNA PCAI protects against cognitive decline induced by hippocampal neuroinflammation via regulating SUZ12: Life Sci, 2020; 253; 117626
45. Ballard C, Jones E, Gauge N, Optimised anaesthesia to reduce postoperative cognitive decline (POCD) in older patients undergoing elective surgery, a randomised controlled trial: PLoS One, 2012; 7(6); e37410 Erratum in: PLoS One. 2012;7(9): 22719840
Figures
Figure 1. Experimental designSprague-Dawley male rats (6–8 weeks old, weighing 200–250 g) were randomly divided into 6 groups, with 6 rats in each group. The adaptation time was 7 days. Acute sleep deprivation lasted for 24 h from day 7, and sevoflurane inhalation lasted for 2 h from day 8. Then, the Y maze learning stage was carried out, and the test stage started on the 9th day. On the 11th day after the end of the Y maze test, the MWM test was started. After all intervention and behavior experiments, rats were killed, and the bilateral hippocampus was removed.Figure 2. Rat survival rate in each groupThe 7-day survival rates of rats in the control group, the 24-h acute sleep deprivation group, the sevoflurane inhalation group, the propofol infusion group, the sleep deprivation combined with sevoflurane group, and the sleep deprivation combined with intravenous anesthesia group were observed. There was no significant difference in the survival rate among groups at different time points (P>0.05).Figure 3. Behavioral changes in Y maze testing in ratsY maze test. (A) Comparison of alternating percentage between automatic Y maze and observer report. The bar graph represents the average value (n=6 for all groups). For comparison, the percentage calculated by the observer is included. Effect of treatment on alternation percentage: The results of the automatic system and the average value of observed values were analyzed independently by one-way ANOVA, and the same results were obtained (* P<0.05 Control vs SD, Control vs Sevo, SD vs SD+Sevo). (B) The number of times that rats in each group entered the new arm within 30 min (* P<0.05 Control vs SD, Control vs Sevo, SD vs SD+Sevo).Figure 4. (A, B) Behavioral changes of MWM test in ratsMorris water maze test. Compared with the Control group, the change of propofol group was small but statistically significant. The acute sleep group and inhalation of sevoflurane group had significantly increased escape latency, and the alternation rate was decreased. Compared with the SD group, sleep deprivation combined with sevoflurane inhalation induced more apparent cognitive impairment, and the difference was statistically significant. However, the cognitive function of the SD combined with propofol group was not significantly different from that of the Control group. (* p<0.05, Control vs SD, Control vs Sevo, SD vs SD+Sevo).Figure 5. Expression of SIK3, PSD-95, and proteins associated with tau modification before administration of inhibitor or agonist in rats(A) Compared with the Control group, the expression level of tau protein in the hippocampus in the SD group was upregulated. Tau protein phosphorylation’s expression level was significantly upregulated, with a decreased acetylation level and an increased phosphorylation/acetylation ratio. The Sevo group exhibited increased SIK3 expression, while other changes were similar to those in the SD group. There was no statistically significant difference between the 2 groups. Compared with the SD group, the expression of SIK3 in the SD+Sevo group was upregulated. The levels of tau and its phosphorylation were increased, and the phosphorylation/acetylation ratio was increased, with statistical significance. In the change of PSD-95, the expression of the SD+Sevo group was significantly reduced. Simultaneously, the SD group and Sevo group also showed a certain degree of expression decline, and the difference was statistically significant. (* P<0.05, Control vs SD, Control vs Sevo, SD vs SD+Sevo). (B) Compared with the Con group, the expressions of SIK3, tau, p-tau were slightly increased, and ace-tau expression was slightly decreased in the SD group. The expression levels of related proteins in the Prop group were unchanged compared with the Con group. SIK3, tau, and p-tau were expressed at reduced levels. The phosphorylation/acetylation ratio was decreased in the SD+Prop group compared with the SD group. Their expression levels were similar to those in the Con group. For PSD-95 expression, SD reduced its expression, and there were no significant differences in PSD-95 expression between Con, Prop, and SD+Prop groups. (* p<0.05, Con vs SD, Con vs Prop, SD vs SD+Prop).Figure 6. (A, B) Expression of SIK3, PSD-95 and proteins associated with tau modification after administration of SIK3 inhibitor in ratsIn the Control group, the expression level of tau protein in the hippocampus of the SD group was upregulated (P<0.05), the expression level of tau protein phosphorylation (Ser404) was significantly upregulated (P<0.05), and the acetylation level was decreased (P<0.05), together with the decrease in postsynaptic PSD95. There was no significant difference in each protein’s levels between the Sevo group and the SD group (P>0.05). The SD+Sevo group showed a stronger trend than the SD group (P<0.05) (n=6/group). (* P<0.05, Con vs SD, Con vs Sevo, SD vs SD+Sevo).Figure 7. (A, B) Expression of SIK3, PSD-95 and proteins associated with tau modification after administration of tau agonist in ratsIntravenous anesthesia with propofol reversed cognitive impairment caused by acute sleep deprivation. In the Control group, the expression level of tau protein in the hippocampus of the SD group was upregulated (P<0.05), the expression level of tau protein phosphorylation (Ser404) was significantly upregulated (P<0.05), and the acetylation level was decreased (P<0.05), together with the decrease in postsynaptic PSD95. There was no significant difference in each protein’s levels between the Prop group and the Control group (P>0.05). The SD+Prop group outperformed the SD group in cognition, and the results were statistically significant (* P<0.05 Con vs SD, Control vs Prop, and SD vs SD+Prop).Figure 8. (A, B) Change in hippocampal dendritic spines before administration of inhibitors or agonistsIn the DG region, the density of dendritic spines decreased significantly in the SD and Sevo groups. There was no noticeable change in the SD group in the basal dendrites of CA 1 pyramidal neurons. However, the density of dendritic spines decreased after inhalation of sevoflurane. This trend also existed in the apical dendritic spines of CA 1. There was no significant difference between the propofol group and the Control group. The SD group density of dendritic spines in the DG and CA 1 area in the SD+Sevo group decreased significantly, suggesting that sleep deprivation and inhalation anesthesia on cognitive function eventually changed synaptic plasticity, thus affecting long-term cognitive function. It is worth noting that the SD+Prop group showed a higher density of dendritic spines. The overall trend was better than that of the sleep deprivation group, which suggests that propofol anesthesia can significantly reverse the cognitive impairment caused by acute sleep deprivation.Figure 9. Change in hippocampal dendritic spines after administration of the SIK3 inhibitor YKL-05-099The changes of bilateral hippocampal neural plasticity in each group were observed by Golgi staining. In the DG region, there was a statistically significant increase in dendritic spine density in the SD and Sevo groups compared to before inhibitor administration. The density of dendritic spines in DG and CA 1 regions in the SD+Sevo group was slightly decreased, suggesting that the effects of sleep deprivation and inhalation anesthesia on cognitive function ultimately changed synaptic plasticity and thus affected long-term cognitive function.Figure 10. Change in hippocampal dendritic spines after administration of Tau modification agonistIn the DG region, dendritic spines’ density was decreased in the SD and Prop groups compared with before. The dendritic spines in DG and CA 1 regions in the SD+Prop group were denser than those in the Control group, suggesting that the effects of sleep deprivation and intravenous anesthesia on cognitive function can eventually change synaptic plasticity, thus affecting long-term cognitive function. Propofol infusion had a definite reversal effect on cognitive dysfunction caused by sleep deprivation. In Press
08 Mar 2024 : Clinical Research
Evaluation of Foot Structure in Preschool Children Based on Body MassMed Sci Monit In Press; DOI: 10.12659/MSM.943765
15 Apr 2024 : Laboratory Research
The Role of Copper-Induced M2 Macrophage Polarization in Protecting Cartilage Matrix in OsteoarthritisMed Sci Monit In Press; DOI: 10.12659/MSM.943738
07 Mar 2024 : Clinical Research
Knowledge of and Attitudes Toward Clinical Trials: A Questionnaire-Based Study of 179 Male Third- and Fourt...Med Sci Monit In Press; DOI: 10.12659/MSM.943468
08 Mar 2024 : Animal Research
Modification of Experimental Model of Necrotizing Enterocolitis (NEC) in Rat Pups by Single Exposure to Hyp...Med Sci Monit In Press; DOI: 10.12659/MSM.943443
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952