26 July 2020: Lab/In Vitro Research
Vitamin D Attenuates Hypoxia-Induced Injury in Rat Primary Neuron Cells through Downregulation of the Dual Oxidase 1 (DUOX1) Gene
Panpan Cui12BD, Yan Wang1AC, Yanzhong Li1AE*, Lei Ge3FDOI: 10.12659/MSM.925350
Med Sci Monit 2020; 26:e925350






Abstract
BACKGROUND: This study aimed to investigate the mechanisms underlying the neuroprotective effects of vitamin D.
MATERIAL AND METHODS: Rat primary neuron cells were incubated under a hypoxia condition [a hypoxic chamber mixed with anaerobic gas (90% N₂, 5% CO₂) and 5% O₂] to induce cell injury. Cell transfection was performed to overexpress or suppress the expression of dual oxidase 1 (DUOX1). The malondialdehyde (MDA) and superoxide dismutase (SOD) levels were detected using a MDA (A003-2) or SOD (A001-1) kit. DUOX1 mRNA levels were detected using RT-PCR. Hypoxia-inducible factor-1α (HIF-1α), DUOX1, vitamin D receptor (VDR), NF-κB protein expressions were determined by western blotting. Cell apoptosis and reactive oxygen species (ROS) were evaluated by flow cytometry.
RESULTS: ROS increased significantly after hypoxic treatment. The expressions of HIF-1α and DUOX1 were significantly increased after hypoxic treatment. Vitamin D could decrease ROS level, apoptotic neuron cells and DUOX1 expression, and increase VDR expression. Downregulation of DUOX1 significantly decreased MDA level and apoptotic percentages of neuron cells, increased SOD level, and counteracted the hypoxia-induced increase of NF-κB signal. Further study showed that overexpression of DUOX1 significantly increased MDA level, ROS level, apoptotic percentages of neuron cells, and NF-κB nuclear signaling, while decreased SOD level. Vitamin D significantly counteracted the effects of DUOX1 overexpression induced injury in rat primary neuron cells.
CONCLUSIONS: Our study indicated that vitamin D may protect neuron cells from hypoxia-induced injury by regulating DUOX1 via the NF-κB signaling pathway.
Keywords: Calcitriol, Cell Adhesion Molecules, Neuronal, Cell Hypoxia, NADPH Dehydrogenase, Dual Oxidases, Hypoxia-Inducible Factor 1, alpha Subunit, Malondialdehyde, Neurons, Neuroprotective Agents, primary cell culture, Receptors, Calcitriol, Vitamin D
Background
Oxygen is an oxidizing agent, the homeostasis of which is necessary to maintain the physiological balance during cell growth and development [1]. Under many pathological conditions, hypoxia interferes with energy metabolism and eventually results in cell death [2]. Neurons in the central nervous system of mammals are vulnerable to hypoxia. The neuronal injury induced by hypoxia is the basis of many neurological disorders, such as stroke. Lack of oxygen in the brain can result in a rapid loss of the electroencephalographic (EEG) activity and evoked potentials, which is a sign of injury [3,4]. It is of great significance to develop promising therapeutic strategies to attenuate hypoxia-induced neuronal injury.
Vitamin D is a steroid hormone that is synthesized in the skin from 7-dehydrocholesterol by ultraviolet light, which is metabolized primarily in the liver and then in the kidney to form calcitriol (1,25(OH)2D3) [5]. Calcitriol is the most bioactive metabolite of vitamin D, which is increasingly recognized for its neuroprotective effects [6]. Thus, vitamin D could not only affect bone mineralization, but also is a potent neurohormone. Previous studies have identified that vitamin D receptors (VDRs) and some enzymes in the vitamin D synthesis pathway are found in the human brain [7,8]. In addition, clinical literature has shown the effects of low vitamin D status on brain injury. For example, Nararsimhan et al. found that supplement of vitamin D significantly improved the outcomes in those ischemic stroke patients after 3 months [9]. Zhou et al. found that lower level of vitamin D was associated with an increased risk of ischemic stroke based on a meta-analysis [10]. It has found that in addition to adults, the status of vitamin D is also associated with neonatal hypoxic ischemic encephalopathy [11–13]. Thus, vitamin D not only plays a critical role in many neurological disorders in adults, but also is important in neuronal development in newborns and children [14]. Nevertheless, the molecular mechanisms underlying the neuroprotective effects of vitamin D remain to be elucidated.
Hypoxia-inducible factor-1 (HIF-1) is a member of the HIF family and is composed of an O2-regulated α subunit and a constitutive β subunit [15]. The transcriptional activity of HIF-1 is induced under continuous hypoxia conditions [16]. Under hypoxia conditions, HIF-1α and HIF-1β form activated HIF-1, which induces the expression of downstream target genes and presents different physiological functions by regulating different target genes, such as angiogenesis, energy metabolism, erythropoiesis, and cell differentiation. In hypoxic-ischemic brain injury, HIF-1 plays a neuropotective role by regulating target gene expression to increase tissue oxygen supply, improve energy metabolism, stimulate angiogenesis and remodeling, and promote nerve regeneration [17,18].
It has been suggested that hypoxic conditions can induce the production of reactive oxygen species (ROS) in neurons, causing lipid peroxidation, and the final product malondialdehyde (MDA) level is increased, while superoxide dismutase (SOD) level is decreased, which can scavenge superoxide free radicals [19,20]. Therefore, MDA and SOD are 2 typical oxidative stress parameters [21]. ROS promotes brain injury via causing damage to lipids, proteins, and nucleic acids, and by activating many redox-sensitive signaling pathways [22,23]. One study demonstrated that cells produce ROS through activating nicotinamide adenine dinucleotide phosphate (NADPH) oxidases that comprise an enzyme family known as NOX/DUOX [24]. It has been reported that NOX2-derived ROS production plays a key role in nerve injury [25]. NOX4 is important for maintaining neuropathic pain after peripheral nerve injury [26]. Dual oxidase (DUOX) enzymes are regulated by some positive and negative signals to adjust the production of ROS to the changing environment [27]. To our best knowledge, there is no direct evidence about the role of DUOX enzymes in neuronal injury.
Thus, in the present study, we intended to investigate whether vitamin D played a neuroprotective role in hypoxia-induced neuronal injury by acting DUOX1 to regulate ROS. Our study may help to understand the mechanisms underlying the neuroprotective effects of vitamin D.
Material and Methods
CELL CULTURE:
Rat primary neuron cells were cultured in a high-glucose Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal bovine serum (FBS) and 1% double antibiotics (penicillin and streptomycin) in a 37°C incubator with 5% CO2.
CONSTRUCTION OF HYPOXIC NERVE INJURY CELL MODEL:
Rat primary neuron cells were set up in a hypoxic group and a normal oxygen group (control). Hypoxic group: cells were cultured in a hypoxic chamber mixed with anaerobic gas (90% N2, 5% CO2) and 5% O2; Control group: cells were cultured in an incubator (95% air, 5% CO2) at 37°C. After 0, 6, 12, and 24 hours of incubation, ROS and HIF-1α was detected to verify that hypoxic nerve injury cell model was successfully constructed.
DUOX1 OVEREXPRESSION AND INTERFERENCE:
DUOX1 (AF542180.1) coding sequences (CDS) were synthesized and validated, and then inserted into pcDNA3.1(+) vector (Addgene, USA) at Hind III/EcoR I sites. DUOX1 overexpression plasmids (oeDUOX1) were extracted using a Plasmid Extraction Kit (Solarbio, Beijing). The sequences of DUOX1 siRNA (siDUOX1) at 3 different sites were synthesized and listed in Table 1.
CELL TRANSFECTION:
Rat primary neuron cells in the logarithmic growth phase were suspended to 1×106 cells/mL after trypsinization. Then 2 mL of suspension was inoculated into 6-well plates for overnight culture at 37°C with 5% CO2. When the cells grew to 60–70% confluency, rat primary neuron cells were transfected with siDUOX1-1, siDUOX1-2, siDUOX1-3 (targeting at 3 different site of DUOX1 gene; MOI=5, 5 μL) and negative control of DUOX1 interference [siNC (MOI=5, 5 μL)], or oeDUOX1 (MOI=5, 5 μL) and empty plasmids (vector, MOI=5, 5 μL) by Lipo2000. After 24 hours of transfection, serum-free transfer solution was replaced by complete medium to culture for 48 hours.
BIOCHEMICAL DETECTION:
After treatment, the cells were collected and the supernatants were obtained to detect MDA and SOD levels using a MDA (A003-2) or SOD (A001-1) kit (Nanjing Jiancheng Biotechnology Research Institute, Jiangsu, China) according to the manufacturer recommendations. Briefly, the kit was mixed with the samples by a vortex mixer. The mouth of the test tube was fastened with a plastic wrap, and a small hole was pierced with a needle, followed by 40 minutes of water bath at 95°C. The absorbance at 532 nm was detected.
REAL-TIME POLYMERASE CHAIN REACTION (RT-PCR) ASSAY:
Total RNA was extracted with TRIzol reagent (1596-026, Invitrogen). After quantification, RNA was reverse transcribed into cDNA by using the Reverse Transcription Kit (#K1622, Fermentas, Lithuania). With cDNA as templates, RT-PCR was carried out on ABI 7300 Real-Time PCR system (Applied Biosystems, ABI-7300, Foster City, CA, USA) with a SYBR Green PCR Kit (#K0223, Thermo Fisher Scientific, Inc., Waltham, MA, USA). Reactions were conducted in triplicate and then normalized to GAPDH. Relative mRNA level was calculated by the method of 2−ΔΔCq. The procedures were as follows: 95°C for 10 minutes; 40 cycles of 95°C for 15 seconds and 60°C for 45 seconds.
The primers sequences were as follows:
WESTERN BLOTTING:
Total proteins were extracted using radioimmunoprecipitation assay (RIPA) buffer (R0010, Solarbio, Beijing, China) and then quantified using a bicinchoninic acid (BCA) quantification kit (PICPI23223, Thermo Fisher Scientific). Then 25 μg proteins were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE; JRDUN Biotechnology Co., Ltd., Shanghai, China) and transferred onto the polyvinylidene fluoride (PVF) membranes (HATF00010, Millipore, Bredford, MA, USA). After blocking in 5% skim milk (BYL40422, BD Biosciences, Franklin Lakes, NJ, USA) for 1 hour at 25°C, the blots were probed with primary antibodies against HIF-1α (1: 400, Ab1, Abcam), DUOX1 (1: 1000, Orb539256, Biorbyt), VDR (1: 1000, Ab109234, Abcam), NF-κB (1: 2000, Ab16502, Abcam), cleaved caspase-3 (1: 1000, AF6311, Affinity), H3 (1: 1000, Ab1791, Abcam), and GAPDH (1: 2000, #5174, Cell Signaling Technology) overnight at 4°C with gentle shaking. Then the blots were washed with TBST 5 to 6 times followed by 1 hour of incubation in secondary antibodies that were labeled with horseradish peroxidase (HRP; 1: 1000, Beyotime, Shanghai, China) at room temperature. After being washed with tris-buffered saline with Tween (TBST) for 5 to 6 times, the blots were developed by 5 minutes incubation with chemiluminescent reagent (WBKLS0100, Millipore). Finally, the protein blots were exposed on an ECL imaging system (Tanon-5200, Tanon, Shanghai, China), and H3 and GAPDH served as endogenous references. Relative protein levels were calculated using ImageJ software (Bethesda, MD, USA).
CELL APOPTOSIS ASSAY:
Flow cytometry analysis was applied to evaluate cell apoptosis. Rat primary neuron cells were collected followed by Annexin V-fluorescein isothiocyanate (FICT)/propidium iodide (PI) double staining (C1052, Beyotime). Then 5×105 to 1×106 cells were resuspended in 195 μL Annexin V-FITC binding buffer, followed by incubated in 5 μL Annexin V-FITC for 15 minutes at 4°C in the dark. Subsequently, the cells were incubated in PI (5 μL) for 5 minutes (4°C, dark). Percentages of apoptotic ZR-75-30 and BT474 cells were evaluated on a flow cytometer by a BD Accuri™ C6 Software (V1.0.264.21, BD Biosciences, USA).
ROS DETECTION:
After treatment, the cell pellets were collected and then resuspended in 1 mL cooled phosphate-buffered saline (PBS). Total 10 mM DCFH-DA probe solution was diluted with serum-free medium at 1: 1000 to obtain a 10 μM staining working solution. The cell samples were obtained and incubated with DCFH-DA probe solution for 20 minutes at 37°C in the dark. Cells and solution were inverted and mixed every 5 minutes. The serum-free medium was washed 3 times, and the samples were detected by a flow cytometry.
STATISTICAL ANALYSIS:
All experiments in this study were conducted thrice independently, and the data were presented as the mean±standard deviation (SD). Statistical testing was performed using the software of GraphPad prism 7.0 (San Diego, CA, USA). ANOVA with Tukey’s multiple comparison post hoc tests was applied for multiple groups.
Results
ROS WAS SIGNIFICANTLY INCREASED IN HYPOXIA-INDUCED INJURY IN RAT PRIMARY NEURON CELLS:
Rat primary neurons were used to construct hypoxic (5% O2) nerve injury cell model. The result showed that compared with the control group (normal oxygen), ROS increased significantly after hypoxic treatment in a time-dependent manner (Figure 1A). As shown in Figure 1B, the expression of DUOX1 mRNA was significantly increased after hypoxic treatment, and the protein expression of HIF-1α and DUOX1 were also significantly increased (Figure 1C). The treatment time of 24 hours was selected for subsequent experiments. These results indicated that ROS increased significantly in hypoxia-induced neuron cells.
VITAMIN D ATTENUATED HYPOXIA-INDUCED INJURY IN RAT PRIMARY NEURON CELLS:
To study the role of vitamin D in hypoxia-induced injury in rat primary neuron cells, the hypoxia-treated neuron cells (24 hours) were treated with different concentrations of vitamin D (0, 50, 100, and 200 nM) for 24 hours. As shown in Figure 2A, ROS decreased significantly with the increase of vitamin D concentration compared with the control group. Additionally, the percentage of apoptotic neuron cells significantly decreased with the increase of vitamin D concentration (Figure 2B). Moreover, the mRNA and protein expression levels of VDR and DUOX1 were detected after vitamin D treatment. The results showed that after vitamin D treatment, the protein expression of VDR significantly increased, while the protein expression of DUOX1 decreased significantly compared with control (Figure 2C–2E). The concentration of 100 nM was selected for subsequent experiments. The results indicated that vitamin D could alleviate hypoxia-induced neuron cell damage, and the effect was significantly enhanced with the increase of vitamin D concentration.
DOWNREGULATION OF DUOX1 SIGNIFICANTLY ATTENUATED HYPOXIA-INDUCED INJURY IN RAT PRIMARY NEURON CELLS:
To further explore the role of DUOX1 in hypoxia-induced injury in rat primary neuron cells, we suppressed the expression of DUOX1 and observed the neuron cell injury condition. First, the interference efficiency of DUOX1 was detected through RT-PCR and western blotting. As shown in Figure 3A and 3B, DUOX1 was downregulated significantly after cell transfection, and siDUOX1-1 and siDUOX1-2 had a better effect to siDUOX1-3. Suppression of DUOX1 significantly decreased MDA level while increased SOD level compared with siNC+ 5%O2 group, which was similar to the effects of vitamin D treatment (Figure 3C, 3D). Additionally, ROS and the apoptotic percentages of neuron cells significantly decreased after DUOX1 was suppressed (Figure 3E, 3F). Furthermore, downregulation of DUOX1 significantly counteracted the hypoxia-induced increase of NF-κB signal in the nucleus (Figure 3G), suggesting that NF-κB signaling may be involved in the regulation of DUOX1 in nerve injury.
VITAMIN D ATTENUATED DUOX1-INDUCED INJURY IN RAT PRIMARY NEURON CELLS:
We further overexpressed DUOX1 in rat primary neuron cells to investigate the role of vitamin D in DUOX1-induced injury in rat primary neuron cells. The overexpression efficiency of DUOX1 is shown in Figure 4A and 4B. The MDA level increased significantly and the SOD level decreased significantly after overexpression of DUOX1, while vitamin D treatment could reverse the effects of overexpressed DUOX1 (Figure 4C, 4D). Moreover, overexpression of DUOX1 significantly increased ROS level, apoptotic percentages of neuron cells, and NF-κB nuclear signaling. Similarly, vitamin D significantly counteracted the effects of DUOX1 overexpression induced injury in rat primary neuron cells (Figure 4E–4G). The results indicated that vitamin D may regulate neuron cell injury through DUOX1.
Discussion
Vitamin D is a vital component of normal neuronal development, deficiency of which is associated with increased vulnerability to neurologic injury. In our study, rat primary neuron cells were incubated under a hypoxia condition to induce cell injury. We found that ROS was significantly increased in hypoxia-induced injury in neuron cells. Vitamin D treatment could attenuate hypoxia-induced injury in neuron cells. Further study revealed that downregulation of DUOX1 played a similar role with vitamin D treatment in hypoxia-induced neuron cells injury. Finally, vitamin D was found to attenuate DUOX1-induced injury in neuron cells, indicating that vitamin D may regulate neuron cell injury through DUOX1.
Intentional production of ROS occurs in many cell types in response to various stimuli [28]. Hypoxic nerve injury can induce the production of a large number of ROS and cause lipid peroxidation reaction to increase the content of end-product MDA. Meanwhile, SOD can remove superoxide radical, thereby decreasing its activity [29,30]. Antioxidant drugs exerts neuroprotective effects by downregulating ROS [31]. In this study, HIF-1α and ROS levels were increased under hypoxia condition, suggesting the neuron cells were damaged. Vitamin D treatment could attenuate hypoxia-induced increase ROS level, suggesting that vitamin D had neuroprotective effects to alleviate hypoxia-induced neuron cell damage.
DUOX1 is a Ca2+-dependent NADPH oxidase, which is responsible for early T cell receptor-stimulated generation of H2O2 [28]. Neurons can express the NADPH oxidase complex [32]. NADPH oxidase is a transmembrane protein complex that reduces oxygen to superoxide by transporting electrons across biological membranes [33]. Study has suggested that nonphagocytic NADPH oxidases play a crucial role in the regulation of physiological and pathophysiological processes [34]. NADPH oxidase activity is a major source of cellular ROS [24]. Our result showed that DUOX1 expression was significantly increased in hypoxia-induced rat neuron cell, while vitamin D treatment could decrease the expression of DUOX1. Interestingly, downregulation of DUOX1 played a similar role with vitamin D treatment in hypoxia-induced neuron cells injury. The results indicated that vitamin D may regulate neuron cell injury through downregulating DUOX1. A previous study of Joseph et al. [35] reports that activation of DUOX1 promotes the activation of NF-κB signaling pathways. In accordance with the aforementioned findings, our study also found that overexpression of DUOX1 significantly increased NF-κB nuclear signaling.
Conclusions
In conclusion, our study indicated that vitamin D may protect neuron cell from hypoxia-induced injury by regulating DUOX1 via the NF-κB signaling pathway. Our results may provide theoretical basis for the application of vitamin D in hypoxia-induced neurological disorders.
Figures
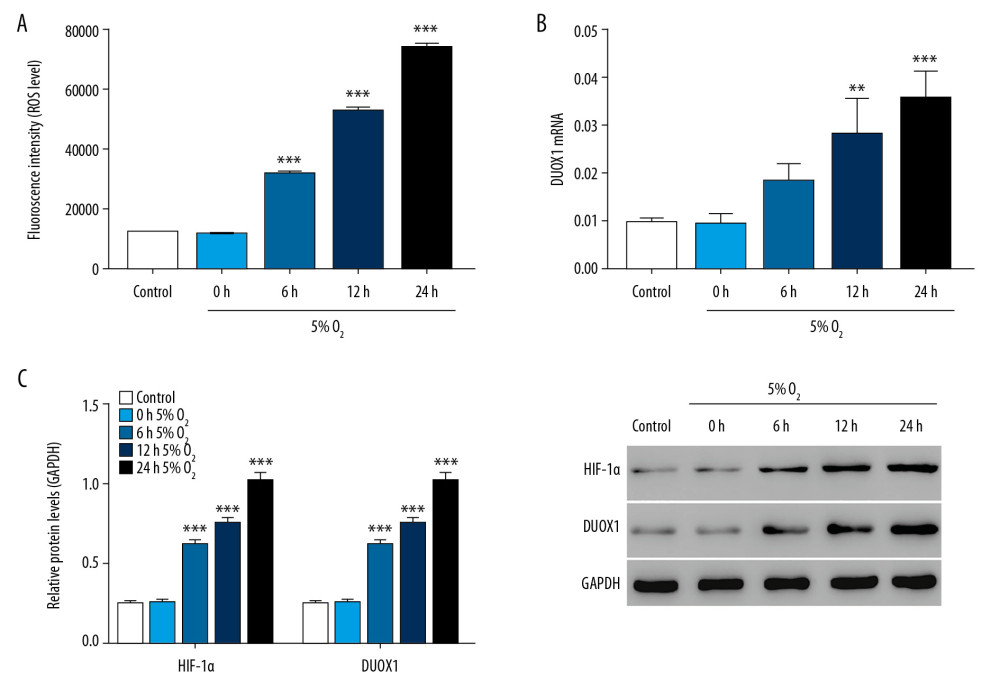 Figure 1. ROS was significantly increased in hypoxia-induced injury in rat primary neuron cells. (A) ROS level was detected by flow cytometry after 0, 6, 12, and 24 hours of hypoxic culture. (B) The mRNA expression of DUOX1 was detected by Q-PCR after 0, 6, 12, and 24 hours of hypoxic culture. (C) The protein expression of HIF-1α and DUOX1 was detected by western blot after 0, 6, 12, and 24 hours of hypoxic culture. * P<0.05, ** P<0.01, *** P<0.001 versus 0 hours 5% O2. ROS – reactive oxygen species; Q-PCR – quantitative polymerase chain reaction; DUOX1 – dual oxidase 1; HIF-1α – hypoxia-inducible factor-1α.
Figure 1. ROS was significantly increased in hypoxia-induced injury in rat primary neuron cells. (A) ROS level was detected by flow cytometry after 0, 6, 12, and 24 hours of hypoxic culture. (B) The mRNA expression of DUOX1 was detected by Q-PCR after 0, 6, 12, and 24 hours of hypoxic culture. (C) The protein expression of HIF-1α and DUOX1 was detected by western blot after 0, 6, 12, and 24 hours of hypoxic culture. * P<0.05, ** P<0.01, *** P<0.001 versus 0 hours 5% O2. ROS – reactive oxygen species; Q-PCR – quantitative polymerase chain reaction; DUOX1 – dual oxidase 1; HIF-1α – hypoxia-inducible factor-1α. 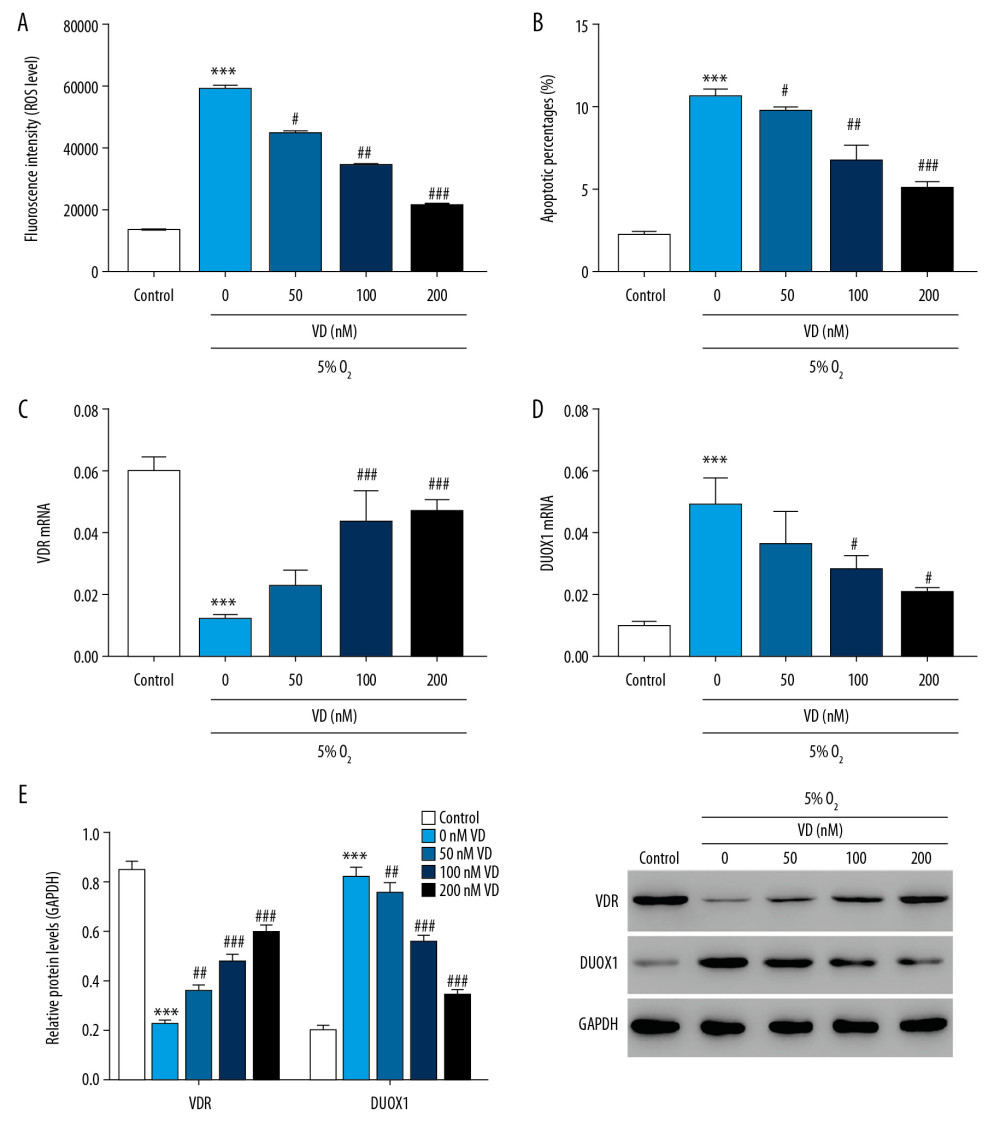 Figure 2. Vitamin D attenuated hypoxia-induced injury in rat primary neuron cells. (A) ROS level was detected by flow cytometry after treatment with different concentrations of vitamin D. (B) Apoptosis was detected by flow cytometry after treatment with different concentrations of vitamin D. (C, D) The mRNA expression of VDR and DUOX1 was detected by Q-PCR after treatment with different concentrations of vitamin D. (E) The protein expression of VDR and DUOX1 was detected by western blot after treatment with different concentrations of vitamin D. *** P<0.001 versus control; # P<0.05, ## P<0.01, ### P<0.001 versus 0 nM vitamin D. Control: rat neuron cells cultured under normoxic conditions. ROS – reactive oxygen species; VDR – vitamin D receptor; DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction;
Figure 2. Vitamin D attenuated hypoxia-induced injury in rat primary neuron cells. (A) ROS level was detected by flow cytometry after treatment with different concentrations of vitamin D. (B) Apoptosis was detected by flow cytometry after treatment with different concentrations of vitamin D. (C, D) The mRNA expression of VDR and DUOX1 was detected by Q-PCR after treatment with different concentrations of vitamin D. (E) The protein expression of VDR and DUOX1 was detected by western blot after treatment with different concentrations of vitamin D. *** P<0.001 versus control; # P<0.05, ## P<0.01, ### P<0.001 versus 0 nM vitamin D. Control: rat neuron cells cultured under normoxic conditions. ROS – reactive oxygen species; VDR – vitamin D receptor; DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction; 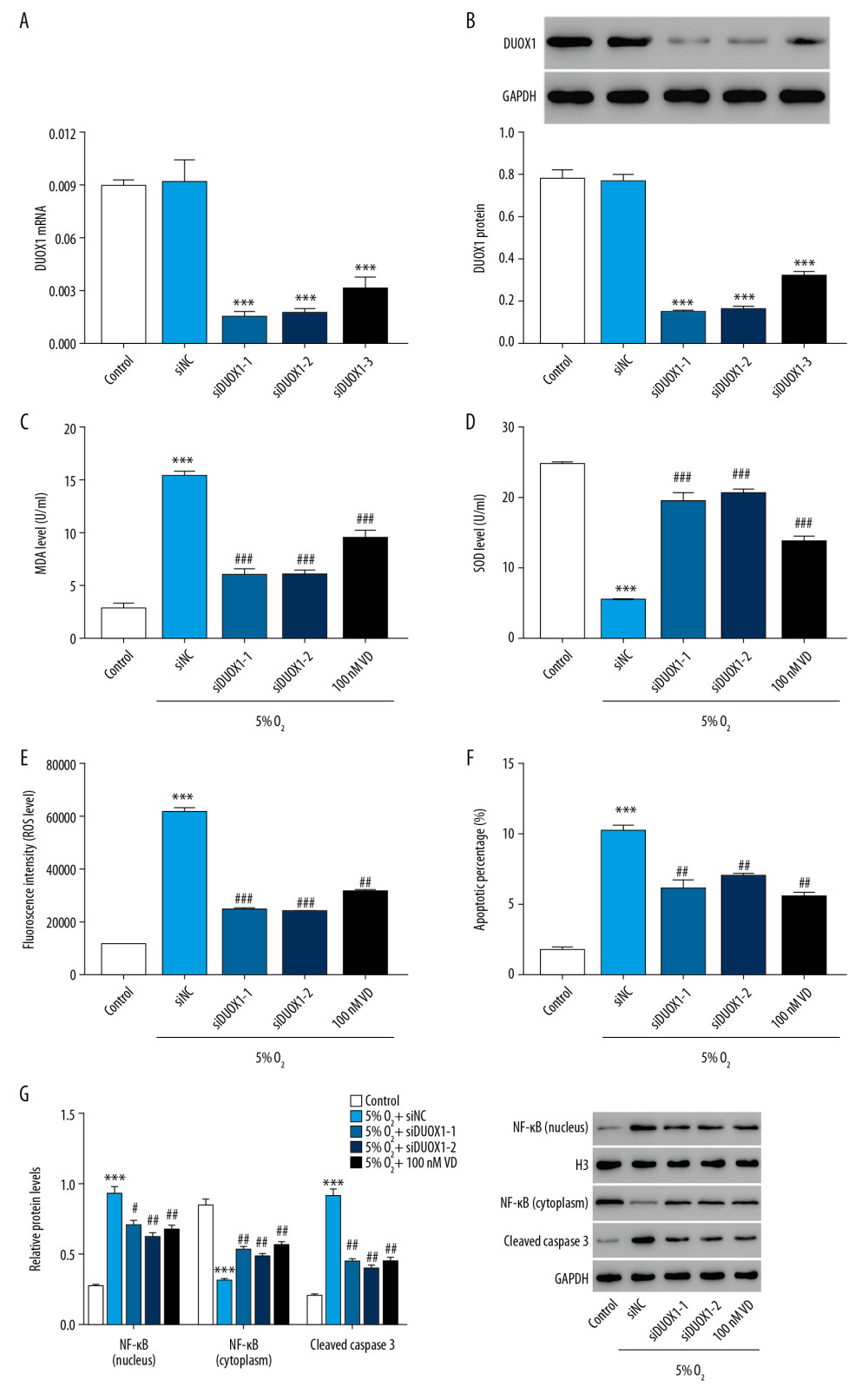 Figure 3. Downregulation of DUOX1 significantly attenuated hypoxia-induced injury in rat primary neuron cells. (A, B) The interference efficiency of DUOX1 was detected by Q-PCR (A) and western blot (B). (C, D) MDA (C) and SOD (D) levels were detected by biochemical detection. (E, F) ROS level (E) and apoptosis (F) were detected by flow cytometry. (G) Protein expression of NF-κB and cleaved caspase-3 was determined by western blot. * P<0.05, ** P<0.01, *** P<0.001 versus siNC or control; # P<0.05, ## P<0.01, ### P<0.001 versus 5% O2+siNC. siNC: Negative control of DUOX1 interference; siDUOX1: DUOX1 interference; Control: rat neuron cells cultured under normoxic conditions. DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction; MDA – malondialdehyde; SOD – superoxide dismutase; ROS – reactive oxygen species.
Figure 3. Downregulation of DUOX1 significantly attenuated hypoxia-induced injury in rat primary neuron cells. (A, B) The interference efficiency of DUOX1 was detected by Q-PCR (A) and western blot (B). (C, D) MDA (C) and SOD (D) levels were detected by biochemical detection. (E, F) ROS level (E) and apoptosis (F) were detected by flow cytometry. (G) Protein expression of NF-κB and cleaved caspase-3 was determined by western blot. * P<0.05, ** P<0.01, *** P<0.001 versus siNC or control; # P<0.05, ## P<0.01, ### P<0.001 versus 5% O2+siNC. siNC: Negative control of DUOX1 interference; siDUOX1: DUOX1 interference; Control: rat neuron cells cultured under normoxic conditions. DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction; MDA – malondialdehyde; SOD – superoxide dismutase; ROS – reactive oxygen species. 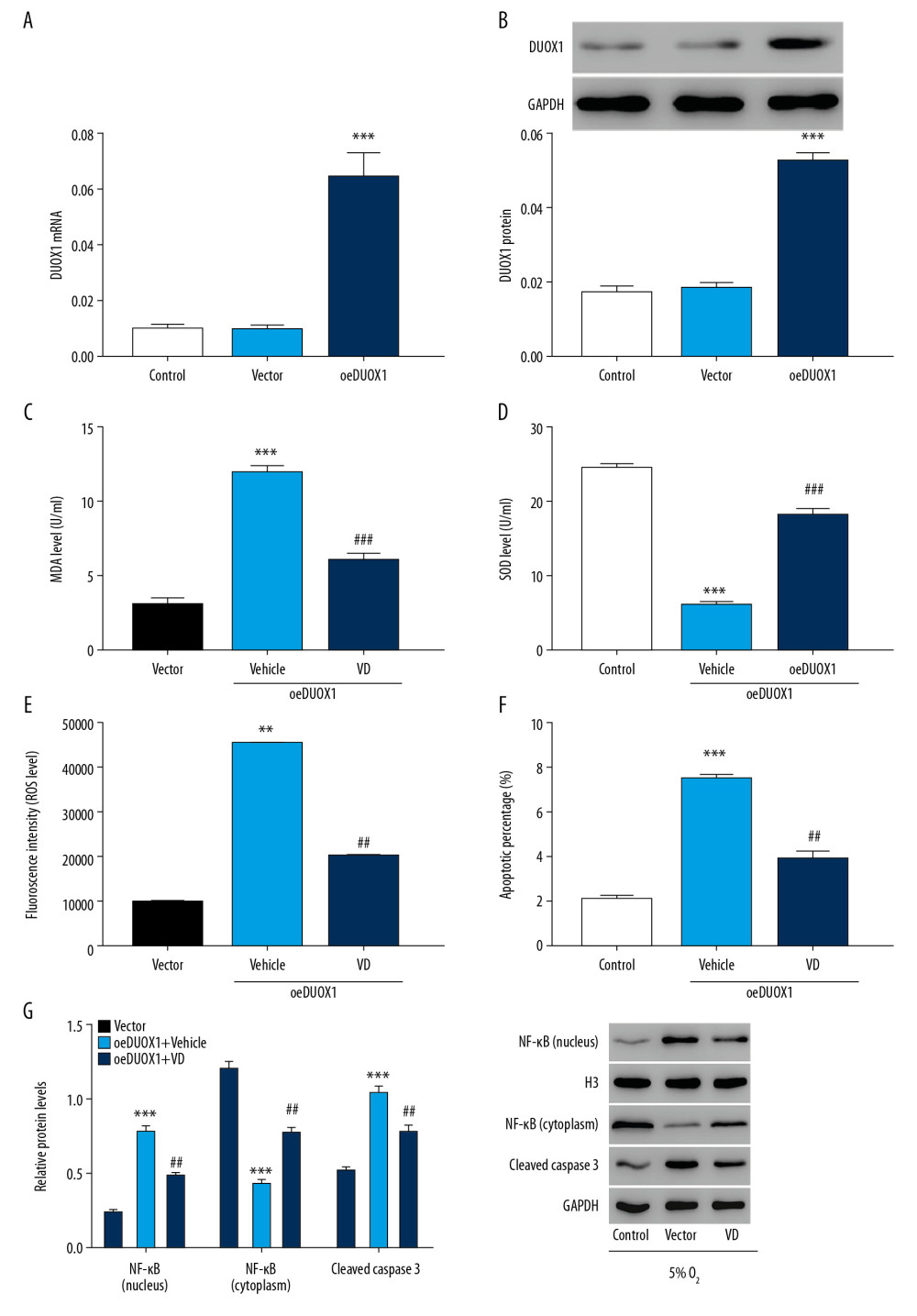 Figure 4. Vitamin D attenuated DUOX1-induced injury in rat primary neuron cells. (A, B) The overexpression efficiency of DUOX1 was detected by Q-PCR (A) and western blot (B). (C, D) MDA (C) and SOD (D) levels were detected by biochemical detection. (E, F) ROS level (E) and apoptosis (F) were detected by flow cytometry. (G) Protein expression of NF-κB and cleaved caspase-3 was determined by western blot. *** P<0.001 versus vector; ## P<0.01, ### P<0.001 versus oeDUOX1+vehicle. Vector: Negative control of DUOX1 overexpression; oeDUOX1: DUOX1 overexpression. DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction; MDA – malondialdehyde; SOD – superoxide dismutase; ROS – reactive oxygen species.
Figure 4. Vitamin D attenuated DUOX1-induced injury in rat primary neuron cells. (A, B) The overexpression efficiency of DUOX1 was detected by Q-PCR (A) and western blot (B). (C, D) MDA (C) and SOD (D) levels were detected by biochemical detection. (E, F) ROS level (E) and apoptosis (F) were detected by flow cytometry. (G) Protein expression of NF-κB and cleaved caspase-3 was determined by western blot. *** P<0.001 versus vector; ## P<0.01, ### P<0.001 versus oeDUOX1+vehicle. Vector: Negative control of DUOX1 overexpression; oeDUOX1: DUOX1 overexpression. DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction; MDA – malondialdehyde; SOD – superoxide dismutase; ROS – reactive oxygen species. 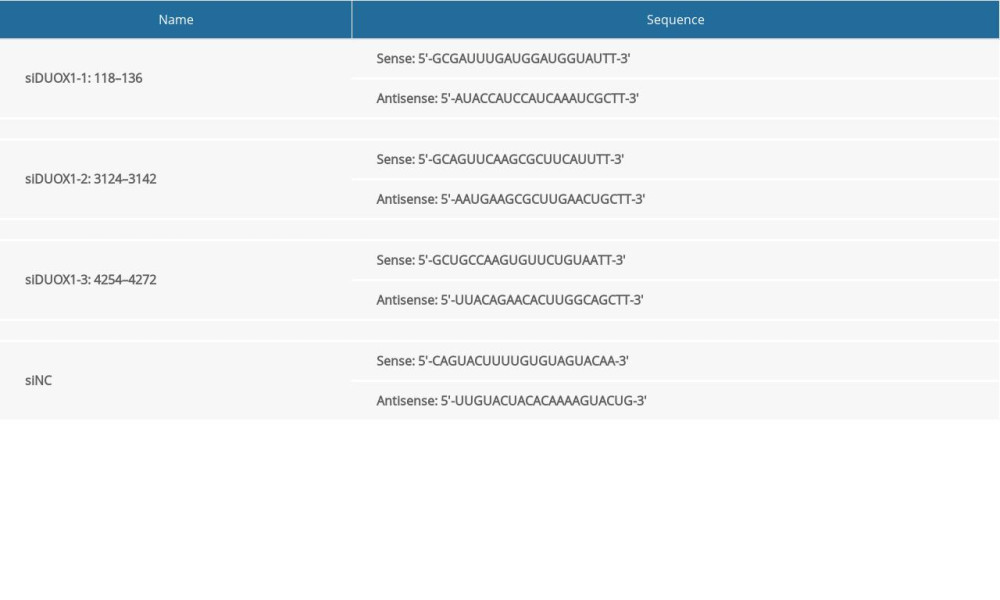
References
1. Samanta D, Prabhakar NR, Semenza GL, Systems biology of oxygen homeostasis: Wires Syst Biol Med, 2017; 9; e1382
2. Saud Alarifi DA, Al Omar Suliman Y, Maqusood Ahamed MAS, Abdulaziz A, Oxidative stress contributes to cobalt oxide nanoparticles-induced cytotoxicity and DNA damage in human hepatocarcinoma cells: Int J Nanomed, 2013; 8; 189-99
3. Haddad G, Jiang C, O2 deprivation in the central nervous system: On mechanisms of neuronal response, differential sensitivity and injury: Prog Neurobiol, 1993; 40; 277-318
4. Fung ML, Role of voltage-gated Na+ channels in hypoxia-induced neuronal injuries: Clin Exp Pharmacol Physiol, 2000; 27; 569-74
5. Won S, Sayeed I, Peterson BL, Vitamin D prevents hypoxia/reoxygenation-induced blood–brain barrier disruption via vitamin D receptor-mediated NF-κB signaling pathways: PLoS One, 2015; 10; e0122821
6. Eelen G, Verlinden L, Van Camp M, The effects of 1α 25-dihydroxyvitamin D3 on the expression of DNA replication genes: J Bone Miner Res, 2004; 19; 133-46
7. Zehnder D, Bland R, Williams MC, Extra renal expression of 25-hydroxy vitamin D3-1α-hydroxylase: J Clin Endocrinol Metab, 2001; 86; 888-94
8. Eyles DW, Smith S, Kinobe R, Distribution of the vitamin D receptor and 1α-hydroxylase in human brain: J Chem Neuroanat, 2005; 29; 21-30
9. Narasimhan S, Balasubramanian P, Role of Vitamin D in the outcome of ischemic stroke – a randomized controlled trial: J Clin Diagn Res, 2017; 11; CC06-10
10. Zhou R, Wang M, Huang H, Lower vitamin D status is associated with an increased risk of ischemic stroke: A systematic review and meta-analysis: Nutrients, 2018; 10; 277
11. Mcginn E, Powers A, Galas M, Neonatal vitamin D status is associated with the severity of brain injury in neonatal hypoxic-ischemic encephalopathy: A pilot study: Neuropediatrics, 2020 [Online ahead of print]
12. Mutlu M, Sariaydin M, Aslan Y, Status of vitamin D, antioxidant enzymes, and antioxidant substances in neonates with neonatal hypoxic-ischemic encephalopathy: J Matern Fetal Neonatal Med, 2015; 29; 2259-63
13. Lowe DW, Hollis BW, Wagner CL, Vitamin D insufficiency in neonatal hypoxic-ischemic encephalopathy: Pediatr Res, 2017; 82; 55-62
14. Stessman LE, Peeples ES, Vitamin D and its role in neonatal hypoxic-ischemic brain injury: Neonatology, 2018; 113; 305-12
15. Wang GL, Jiang B-H, Rue EA, Semenza GL, Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension: Proc Natl Acad Sci USA, 1995; 92; 5510-14
16. Coleman ML, Ratcliffe PJ, Oxygen sensing and hypoxia-induced responses: Essays Biochem, 2007; 43; 1-15
17. Prabhakar NR, Kumar GK, Nanduri J, Intermittent hypoxia augments acute hypoxic sensing via HIF-mediated ROS: Respir Physiol Neurobiol, 2010; 174; 230-34
18. Fan X, Heijnen CJ, van der Kooij MA, The role and regulation of hypoxia-inducible factor-1α expression in brain development and neonatal hypoxic – ischemic brain injury: Brain Res Rev, 2009; 62; 99-108
19. Uttara B, Singh AV, Zamboni P, Mahajan R, Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options: Cur Neuropharmacol, 2009; 7; 65-74
20. Martinez MA, Rodriguez J, Lopeztorres B, Oxidative stress and related gene expression effects of cyfluthrin in human neuroblastoma SH-SY5Y cells: Protective effect of melatonin: Environ Res, 2019; 177; 108579
21. Omidi A, Namazi F, Jabire S, The effects of starvation and refeeding on oxidative stress parameters (MDA, SOD, GPx), lipid profile, thyroid hormones and thyroid histopathology in male Wistar rats: Int Arch Med, 2016; 9; 238
22. Pun PB, Lu J, Moochhala S, Involvement of ROS in BBB dysfunction: Free Radic Res, 2009; 43; 348-64
23. Wang L-F, Li X, Gao Y-B, Activation of VEGF/Flk-1-ERK pathway induced blood–brain barrier injury after microwave exposure: Mol Neurobiol, 2015; 52; 478-91
24. Bedard K, Krause K-H, The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology: Physiol Rev, 2007; 87; 245-313
25. Kim D, You B, Jo E-K, NADPH oxidase 2-derived reactive oxygen species in spinal cord microglia contribute to peripheral nerve injury-induced neuropathic pain: Proc Natl Acad Sci USA, 2010; 107; 14851-56
26. Kallenborn-Gerhardt W, Schröder K, Del Turco D, NADPH oxidase-4 maintains neuropathic pain after peripheral nerve injury: J Neurosci, 2012; 32; 10136-45
27. Leulier F, Royet J, Maintaining immune homeostasis in fly gut: Nat Immunol, 2009; 10; 936-38
28. Kwon J, Shatynski KE, Chen H, The nonphagocytic NADPH oxidase Duox1 mediates a positive feedback loop during T cell receptor signaling: Sci Signal, 2010; 3; ra59
29. Martínez M-A, Rodríguez J-L, Lopez-Torres B, Oxidative stress and related gene expression effects of cyfluthrin in human neuroblastoma SH-SY5Y cells: protective effect of melatonin: Environ Res, 2019; 177; 108579
30. Zhao H-B, Lin S-Q, Liu J-H, Lin Z-B, Polysaccharide extract isolated from Ganoderma lucidum protects rat cerebral cortical neurons from hypoxia/reoxygenation injury: J Pharmacol Sci, 2004; 95; 294-98
31. Tataranno ML, Perrone S, Longini M, Buonocore G, New antioxidant drugs for neonatal brain injury: Oxid Med Cell Longev, 2015; 2015 108251
32. Chéret C, Gervais A, Lelli A, Neurotoxic activation of microglia is promoted by a nox1-dependent NADPH oxidase: J Neurosci, 2008; 28; 12039-51
33. Lu Q, Wainwright MS, Harris VA, Increased NADPH oxidase-derived superoxide is involved in the neuronal cell death induced by hypoxia – ischemia in neonatal hippocampal slice cultures: Free Radic Biol Med, 2012; 53; 1139-51
34. Mittal M, Roth M, König P, Hypoxia-dependent regulation of nonphagocytic NADPH oxidase subunit NOX4 in the pulmonary vasculature: Circ Res, 2007; 101; 258-67
35. Joseph T, Look D, Ferkol T, NF-κB activation and sustained IL-8 gene expression in primary cultures of cystic fibrosis airway epithelial cells stimulated with Pseudomonas aeruginosa: Am J Physiol Lung Cell Mol Physiol, 2005; 288; L471-79
Figures
Figure 1. ROS was significantly increased in hypoxia-induced injury in rat primary neuron cells. (A) ROS level was detected by flow cytometry after 0, 6, 12, and 24 hours of hypoxic culture. (B) The mRNA expression of DUOX1 was detected by Q-PCR after 0, 6, 12, and 24 hours of hypoxic culture. (C) The protein expression of HIF-1α and DUOX1 was detected by western blot after 0, 6, 12, and 24 hours of hypoxic culture. * P<0.05, ** P<0.01, *** P<0.001 versus 0 hours 5% O2. ROS – reactive oxygen species; Q-PCR – quantitative polymerase chain reaction; DUOX1 – dual oxidase 1; HIF-1α – hypoxia-inducible factor-1α.Figure 2. Vitamin D attenuated hypoxia-induced injury in rat primary neuron cells. (A) ROS level was detected by flow cytometry after treatment with different concentrations of vitamin D. (B) Apoptosis was detected by flow cytometry after treatment with different concentrations of vitamin D. (C, D) The mRNA expression of VDR and DUOX1 was detected by Q-PCR after treatment with different concentrations of vitamin D. (E) The protein expression of VDR and DUOX1 was detected by western blot after treatment with different concentrations of vitamin D. *** P<0.001 versus control; # P<0.05, ## P<0.01, ### P<0.001 versus 0 nM vitamin D. Control: rat neuron cells cultured under normoxic conditions. ROS – reactive oxygen species; VDR – vitamin D receptor; DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction;Figure 3. Downregulation of DUOX1 significantly attenuated hypoxia-induced injury in rat primary neuron cells. (A, B) The interference efficiency of DUOX1 was detected by Q-PCR (A) and western blot (B). (C, D) MDA (C) and SOD (D) levels were detected by biochemical detection. (E, F) ROS level (E) and apoptosis (F) were detected by flow cytometry. (G) Protein expression of NF-κB and cleaved caspase-3 was determined by western blot. * P<0.05, ** P<0.01, *** P<0.001 versus siNC or control; # P<0.05, ## P<0.01, ### P<0.001 versus 5% O2+siNC. siNC: Negative control of DUOX1 interference; siDUOX1: DUOX1 interference; Control: rat neuron cells cultured under normoxic conditions. DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction; MDA – malondialdehyde; SOD – superoxide dismutase; ROS – reactive oxygen species.Figure 4. Vitamin D attenuated DUOX1-induced injury in rat primary neuron cells. (A, B) The overexpression efficiency of DUOX1 was detected by Q-PCR (A) and western blot (B). (C, D) MDA (C) and SOD (D) levels were detected by biochemical detection. (E, F) ROS level (E) and apoptosis (F) were detected by flow cytometry. (G) Protein expression of NF-κB and cleaved caspase-3 was determined by western blot. *** P<0.001 versus vector; ## P<0.01, ### P<0.001 versus oeDUOX1+vehicle. Vector: Negative control of DUOX1 overexpression; oeDUOX1: DUOX1 overexpression. DUOX1 – dual oxidase 1; Q-PCR – quantitative polymerase chain reaction; MDA – malondialdehyde; SOD – superoxide dismutase; ROS – reactive oxygen species. In Press
15 Apr 2024 : Laboratory Research
The Role of Copper-Induced M2 Macrophage Polarization in Protecting Cartilage Matrix in OsteoarthritisMed Sci Monit In Press; DOI: 10.12659/MSM.943738
07 Mar 2024 : Clinical Research
Knowledge of and Attitudes Toward Clinical Trials: A Questionnaire-Based Study of 179 Male Third- and Fourt...Med Sci Monit In Press; DOI: 10.12659/MSM.943468
08 Mar 2024 : Animal Research
Modification of Experimental Model of Necrotizing Enterocolitis (NEC) in Rat Pups by Single Exposure to Hyp...Med Sci Monit In Press; DOI: 10.12659/MSM.943443
18 Apr 2024 : Clinical Research
Comparative Analysis of Open and Closed Sphincterotomy for the Treatment of Chronic Anal Fissure: Safety an...Med Sci Monit In Press; DOI: 10.12659/MSM.944127
Most Viewed Current Articles
17 Jan 2024 : Review article
Vaccination Guidelines for Pregnant Women: Addressing COVID-19 and the Omicron VariantDOI :10.12659/MSM.942799
Med Sci Monit 2024; 30:e942799
14 Dec 2022 : Clinical Research
Prevalence and Variability of Allergen-Specific Immunoglobulin E in Patients with Elevated Tryptase LevelsDOI :10.12659/MSM.937990
Med Sci Monit 2022; 28:e937990
16 May 2023 : Clinical Research
Electrophysiological Testing for an Auditory Processing Disorder and Reading Performance in 54 School Stude...DOI :10.12659/MSM.940387
Med Sci Monit 2023; 29:e940387
01 Jan 2022 : Editorial
Editorial: Current Status of Oral Antiviral Drug Treatments for SARS-CoV-2 Infection in Non-Hospitalized Pa...DOI :10.12659/MSM.935952
Med Sci Monit 2022; 28:e935952